17
LA MEMORIA

LA MEMORIA
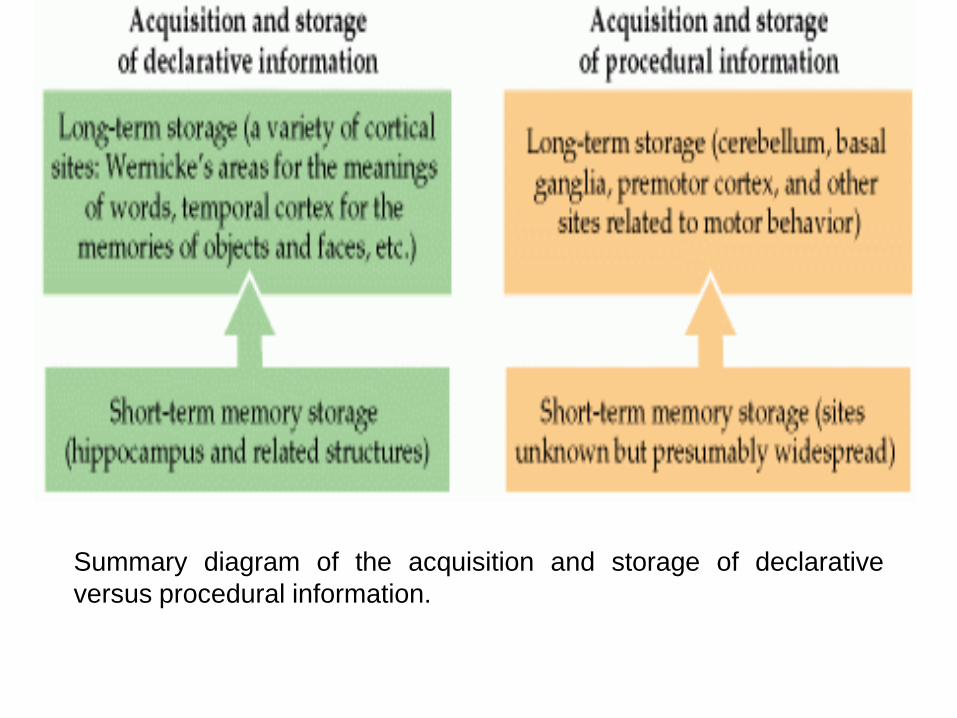
Summary diagram of the acquisition and storage of declarative
versus procedural information.


Principali regioni encefaliche con funzione regolatrice



BASI MOLECOLARI
DEI PROCESSI DI MEMORIZZAZIONE
E DELLA MEMORIA
Fenomeni apparentemente opposti
“di apprendimento” (abituazione e
sensibilizzazione) possono essere
rilevati, nei tessuti eccitabili, dopo
stimoli reiterati.
Se di breve durata (qualche decina
di secondi o pochi minuti), essi
possono avvenire anche all’interno
di un circuito monosinaptico, per
inattivazione di tappe essenziali
delle sinapsi o per squilibrio
temporaneo dell’ambiente ionico.

In sistemi più complessi intervengono invece afferenze da parte di
interneuroni “terzi”, che modificano la stimolabilità del bottone
presinaptico o dei recettori postsinaptici.

Effetti di maggiore durata (ore)
avvengono invece a seguito
dell’ingresso nel bottone
presinaptico (ad opera della
stimolazione reiterata) di “ioni
modificatori” oppure per la
formazione di “secondi
messaggeri” in grado di
modificare l’attività di alcuni
enzimi o delle proteine recettoriali
deputate all’interazione con i
neurotrasmettitori. Uno dei
principali ioni modificatori è lo
ione calcio (Ca2+), mentre uno dei
principali secondi messaggeri è
l’adenosinmonofosfato-3’-5’ciclico
(cAMP), che attiva enzimi (le
“protein-chinasi”) che aggiungono
gruppi fosfato ad alcuni
aminoacidi delle sequenze
proteiche.

Tra i substrati delle proteinchinasi attivate dal
cAMP vi è anche la proteina nucleare CREB
(proteina che si lega ad un elemento genico
responsivo al cAMP) che, previa interazione
con lo stesso cAMP, interagisce con il DNA,
facendo sì che siano espressi geni che
codificano per altri enzimi modificatori di
proteine e che vi sia la biosintesi di nuove
sinapsi e/o nuove vie nervose. I processi di
fosforilazione o defosforilazione della proteina
CREB possono essere anche regolati dagli ioni
Ca++, con effetti però diversi, spesso opposti, a
seconda della loro concentrazione.


Analoghi processi elementari si sono
dimostrati in grado di “spiegare” effetti
di “memoria” nei circuiti nervosi
dell’ippocampo degli animali superiori.
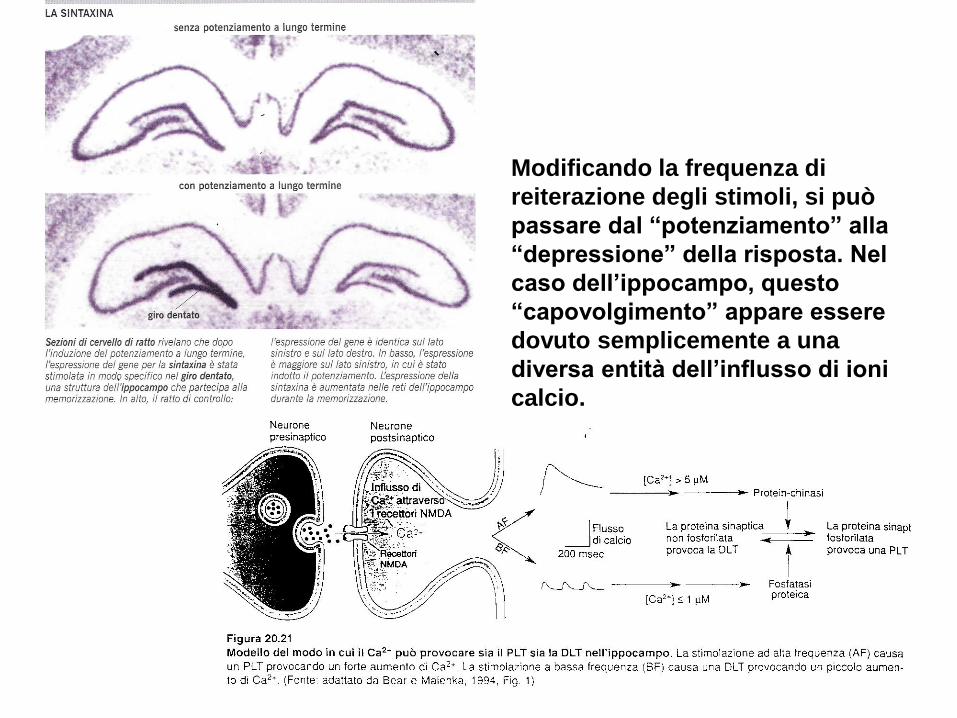
Modificando la frequenza di
reiterazione degli stimoli, si può
passare dal “potenziamento” alla
“depressione” della risposta. Nel
caso dell’ippocampo, questo
“capovolgimento” appare essere
dovuto semplicemente a una
diversa entità dell’influsso di ioni
calcio.
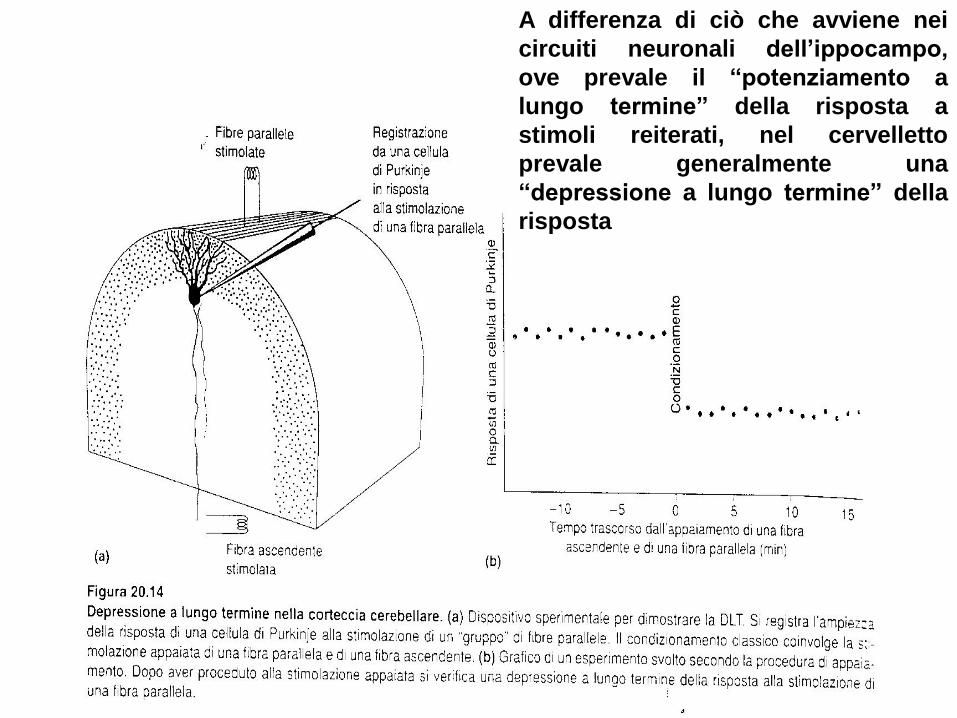
A differenza di ciò che avviene nei
circuiti neuronali dell’ippocampo,
ove prevale il “potenziamento a
lungo termine” della risposta a
stimoli reiterati, nel cervelletto
prevale generalmente una
“depressione a lungo termine” della
risposta

Anche nei circuiti cerebellari si possono tuttavia evidenziare fenomeni di potenziamento.

NEUROTROFINE E NEO-NEUROGENESI Le neurotrofine (NGF, BDNF,
NT3, NT4: peptidi prodotti e
secreti da vari neuroni)
possono, interagendo con
specifici recettori presenti sulle
membrane plasmatiche
neuronali, stimolare la
biogenesi di dendriti e di
nuove sinapsi e assicurare la
sopravvivenza dei neuroni. Nel
giro dentato dell’ippocampo è
stata addirittura evidenziata la
formazione di nuovi neuroni a
partire da cellule staminali. Si
possono avere però effetti
opposti (apoptosi) se, a
seguito di stimoli impropri, vi è
interazione tra i recettori ed i
precursori (pro-neurotrofine)
delle neurotrofine mature. Ciò
si riflette, ovviamente, sulla
memoria.

Nella Drosophila sono stati caratterizzati vari geni
coinvolti nei processi di “memoria”. Questi geni
son presenti, con strutture analoghe o addirittura
identiche, anche negli animali superiori, uomo
compreso.
La maggior parte di questi
geni codificano per la
biosintesi di proteine
coinvolte in tappe
elementari dei processi di
memorizzazione.

















