70
L’acqua Le proprietà chimico-fisiche e gli effetti nei sistemi biologici Il potenziale idrico e il movimento dell’acqua nelle cellule vegetali

L’acqua
�Le proprietà chimico-fisiche e gli effetti nei sistemi biologici
�Il potenziale idrico e il movimento dell’acqua nelle cellule vegetali

N. Rascio – Elementi di Fisiologia Vegetale - Capitolo 2
L’acqua si muove dal terreno all’atmosfera
attraverso la pianta:
� l’acqua presente nel terreno viene
assorbita dalle radici
� viaggia nei vasi conduttori fino alle foglie
� diffonde nell’atmosfera attraverso gli
stomi
Il continuum suolo-pianta-atmosfera

N. Rascio – Elementi di Fisiologia Vegetale - Capitolo 2
Gli stomi sono costituiti da duecellule specializzate che delimitanoun’apertura.
I vasi conduttori sonoformati da cellulesovrapposte in mododa formare dei tubiper il trasportodell’acqua.
Le piante controllano il livello di apertura stomatica e modellano la morfologia dei vasi conduttori in funzione delle condizioni ambientali in modo da
ottimizzare la perdita di acqua in condizioni di stress ambientale
Ghuge et al., (2015) Plant Physiol. 168:690-707..Ghuge et al., (2015) Plant Signal Behav. 10: e1073872.

L’assorbimento della CO2 è accoppiato alla perdita d’acqua sotto forma di vapore
L’acqua è perciò un fattore limitante per le piante che la utilizzano in quantità enormi
L’ACQUA E LE PIANTE
Le piante terrestri assorbono la CO2 dall’atmosfera attraverso gli stomi
Poiché il gradiente di concentrazione per la perdita di H2O dalle foglie è molto più grande di quello per l’assorbimento della CO2 (l’aria esterna è invariabilmente più secca di quella negli spazi intercellulari) a stomi aperti l’H2O evapora verso l’esterno della foglia con un conseguente pericolo di disidratazione

L'ordine di grandezza delle quantità traspirate è elevato: una normale pianta di mais traspira, nella sua vita di circa sei mesi, circa 200 litri d'acqua, l'equivalente del contenuto di un barile
500 grammi di H 2O traspirata per ogni grammo di CO 2 incorporata
H2O assorbita
�97% traspirazione
�3% crescita e metabolismo

Variazioni del contenuto idrico possono causare danni consistenti
a numerosi processi cellularie limitano fortemente la
produttività degli ecosistemi e delle specie coltivate
Il bilancio idrico deve sempre essere in positivo
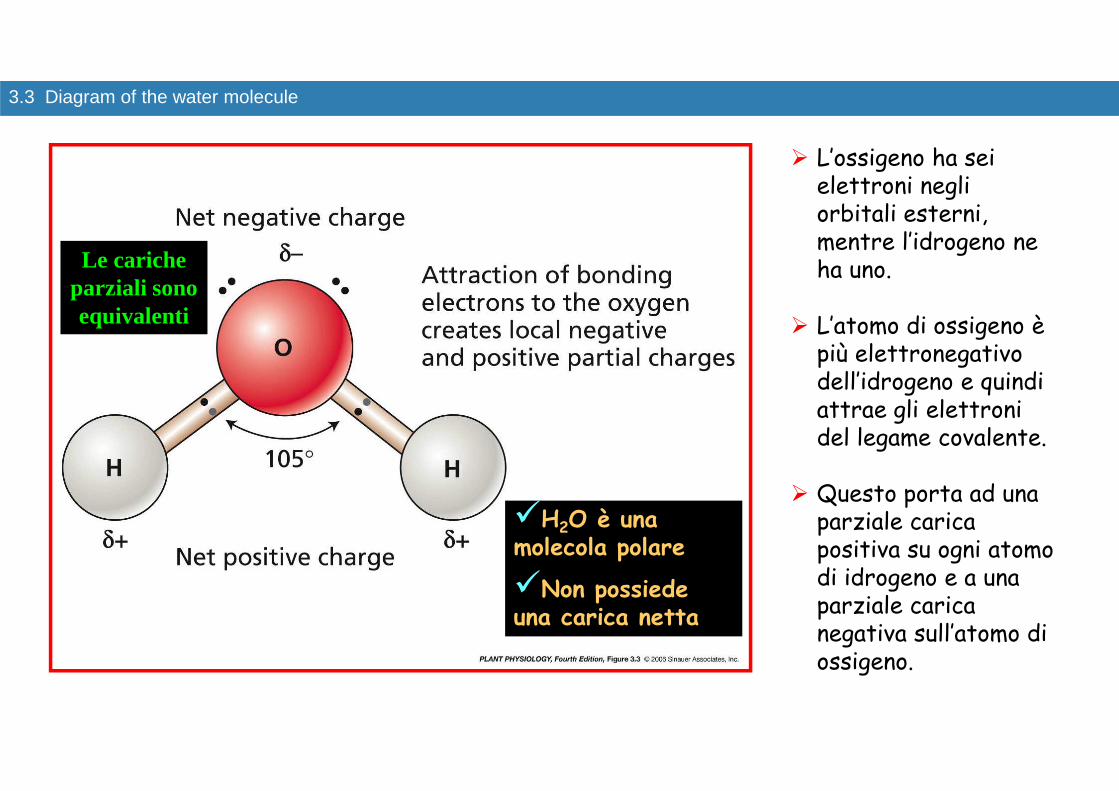
3.3 Diagram of the water molecule
� L’ossigeno ha sei elettroni negli orbitali esterni, mentre l’idrogeno ne ha uno.
� L’atomo di ossigeno è più elettronegativo dell’idrogeno e quindi attrae gli elettroni del legame covalente.
� Questo porta ad una parziale carica positiva su ogni atomo di idrogeno e a una parziale carica negativa sull’atomo di ossigeno.
Le cariche parziali sono equivalenti
�H2O è una molecola polare
�Non possiede una carica netta
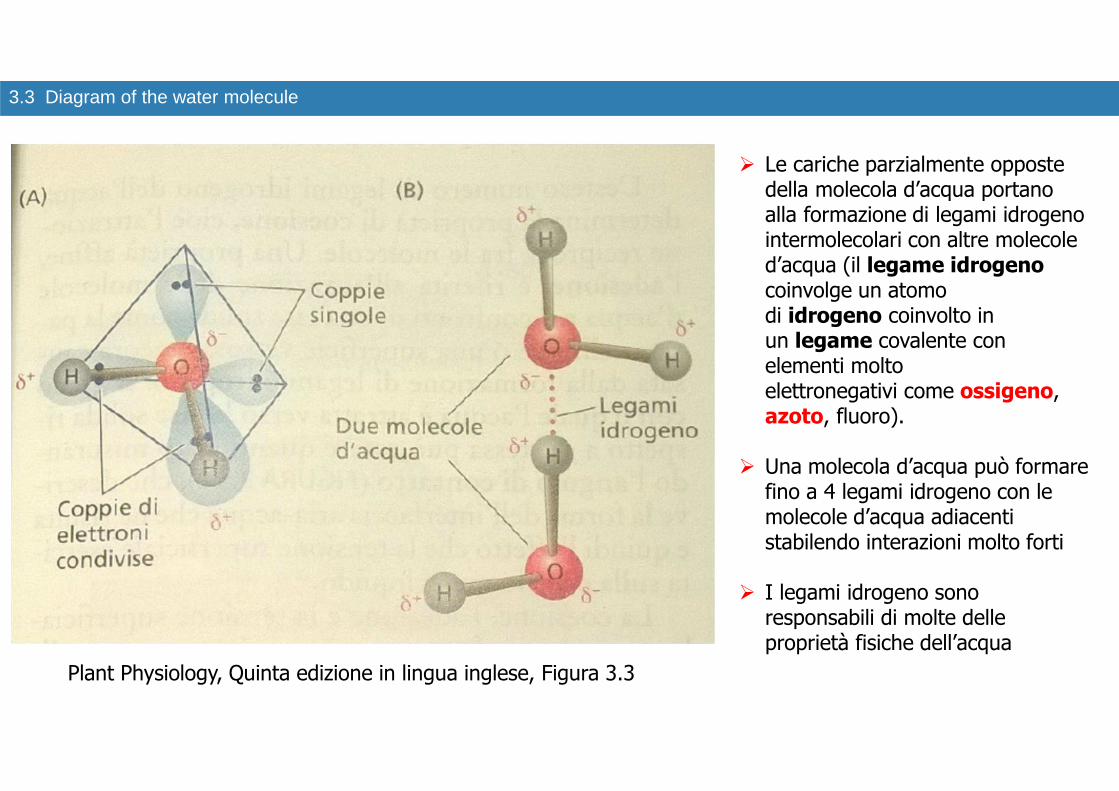
3.3 Diagram of the water molecule
Plant Physiology, Quinta edizione in lingua inglese, Figura 3.3
� Le cariche parzialmente opposte della molecola d’acqua portano alla formazione di legami idrogeno intermolecolari con altre molecole d’acqua (il legame idrogenocoinvolge un atomo di idrogeno coinvolto in un legame covalente con elementi molto elettronegativi come ossigeno, azoto, fluoro).
� Una molecola d’acqua può formare fino a 4 legami idrogeno con le molecole d’acqua adiacenti stabilendo interazioni molto forti
� I legami idrogeno sono responsabili di molte delle proprietà fisiche dell’acqua

Il prodotto ionico dell’acqua
2 H2O ↔ H3O+ + OH-
Lo ione idrogeno tende a dissociare dall’atomo di
ossigeno a cui è legato covalentemente e a saltare
sull’atomo di ossigeno della molecola d’acqua adiacente
a cui è legato da un ponte idrogeno.

Il prodotto ionico dell’acqua
2 H2O ↔ H3O+ + OH-
[H+] [OH-]
Keq =
[H2O]
[H2O] = 55.5 M
� 1000/18 = 55,5 M
� 1000 sono i grammi di H2O
contenuti in un litro di acqua
� 18 è il peso molecolare dell’acqua
La concentrazione dell’acqua non è modificata significativamente dalla sua deboleionizzazione in quanto la concentrazione dell’acqua è molto alta e le concentrazioni degli ioniH+ e OH- sono molto basse (1 x 10-7 M a 25 °C).
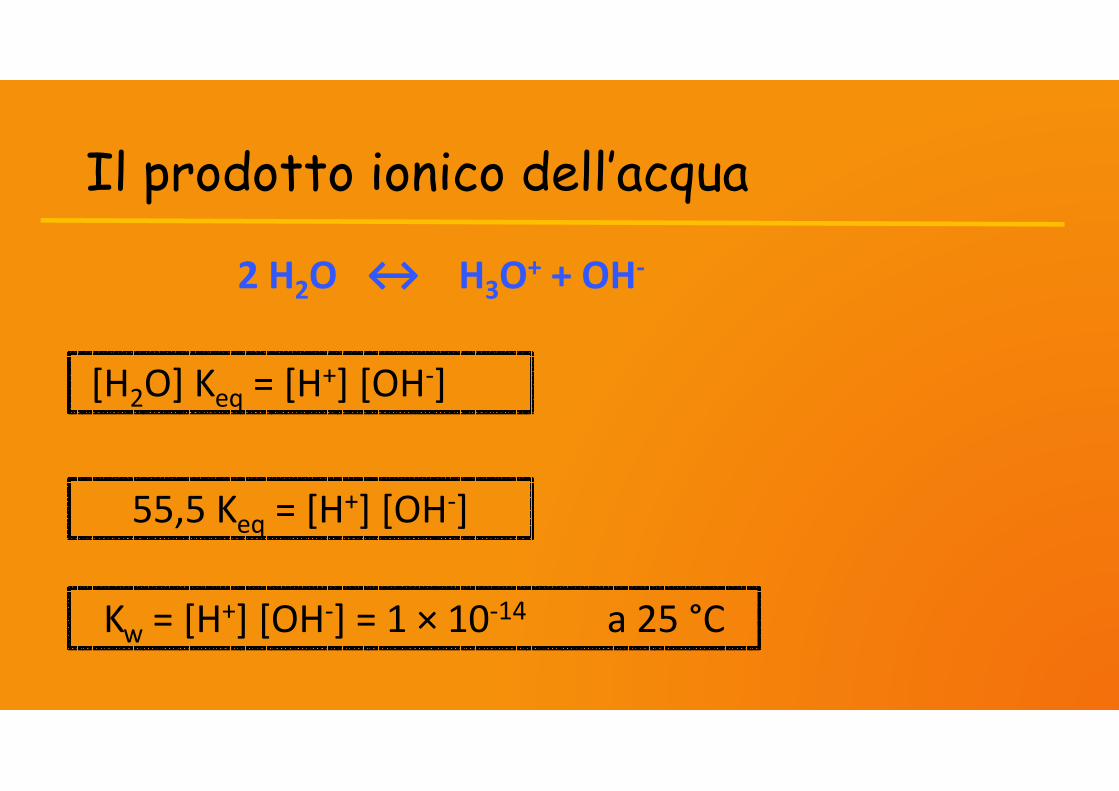
Il prodotto ionico dell’acqua
2 H2O ↔ H3O+ + OH-
[H2O] Keq = [H+] [OH-]
55,5 Keq = [H+] [OH-]
Kw = [H+] [OH-] = 1 × 10-14 a 25 °C

Il prodotto ionico dell’acqua: la scala del pH
2 H2O ↔ H3O+ + OH-
pH = log10 1/[H+] = -log10 [H+]
[H+] = [OH-] = 1 x 10-7 M a 25 °C in soluzione neutra
pH = log10 1/1 x 10-7 M = 7

� L’ attrazione reciproca fra molecole di H2O e l’esteso numero di legami idrogeno che ne conseguono, determinano le proprietà coesivedell’H2O
�Queste proprietà conferiscono all’acqua un importante ruolo nei processi di termoregolazione negli organismi viventi
Le proprietà coesive dell’acqua

Le proprietà coesive dell’acqua : La termoregolazione negli organismi viventi
Alto calore specifico È l’energia di calore richiesta per innalzare la temperatura di 1 grammo di H2O pura di 1 °C (con l’aumentare della temperatura le molecole vibrano più velocemente). Questo determina un elevato punto di ebollizione.
Riduce le fluttuazioni dannose della temperatura
Alto calore latente di evaporazioneÈ l’energia richiesta per separare le molecole di 1 grammo di H2O pura dalla fase liquida e spostarle nella fase gassosa a temperatura costante (44 kJ mol-1 a 25°C)
Importante per il raffreddamento durante l’evaporazione del sudore la traspirazione nelle piante

La formazione del legame idrogeno conferisce all’acqua la capacità di solvatare e quindi
di sciogliere in essa sia ioni (con carica netta), sia molecole organiche (ad esempio
zuccheri) e macromolecole (ad es. proteine) contenenti gruppi funzionali (quali ad es.
gruppi –OH e -NH2) in grado di partecipare alla formazione dei legami idrogeno con le
molecole d’acqua stesse
� Le cellule sono costituite circa dall’80 % di acqua
� Le deboli interazioni causate dal legame idrogeno sono fondamentali per i processi cellulari, perché possono facilmente formarsi e rompersi in condizioni fisiologiche
� Inoltre legami idrogeno possono formarsi anche fra l’H2O e altre molecole che contengono un atomo elettronegativo
L’acqua quale solvente per le reazioni biochimiche

Di Taxman - http://bio.winona.edu/berg/ILLUST/Na+H2O.gif, Pubblico dominio,
https://commons.wikimedia.org/w/index.php?curid=3648176
http://www.chimicare.org/curiosita/wp-
content/uploads/2015/01/solvatazione-sodio-cloruro.jpg
L’acqua e la solvatazione delle molecole in soluzione

https://www.google.it/search?q=acqua+solvatazione&source=lnms&tbm=isch&sa=X&ved=0ahUKEwjM4rPhi_zWAhXBchQKHTQCCDYQ_AUICygC&biw
=1240&bih=532#imgrc=5imChoWnmxJ7GM:
La termodinamica della solubilizzazione

Come fa l’acqua a scorrere verso l’alto quando si
muove verso la cima di un grande albero?

IL POTENZIALE ELETTROCHIMICO
Il potenziale elettrochimico
di un soluto è definito come la somma del potenziale chimico in condizioni standard e dei potenziali di concentrazione, elettrico, idrostatico e di gravità
E’ l’energia disponibile per compiere lavoro
μ = μ0 + RTln a + zFE + PV + mgh
μ0 = potenziale chimico in condizioni standardRTln a = componente della concentrazione (attività)zFE = componente del potenziale elettricoPV = componente della pressione idrostatica mgh = componente della forza di gravità

IL POTENZIALE CHIMICO DELL’ACQUA
�μ0= potenziale standard (per convenzione = 0) riferito all’acqua pura a pressione atmosferica e alla T considerata
�a = attività dell’acqua (per H2O pura = 1 per convenzione)
�V = volume di una mole d’acqua = 18.03 cm3/mol ovvero 18x10-6 m3 mol-1
�m = massa dell’acqua =18
L’acqua è una sostanza polare ma non carica per cui si elimina il componente del potenziale elettrico
μ = μ0 + RTln a + zFE + PV + mgh
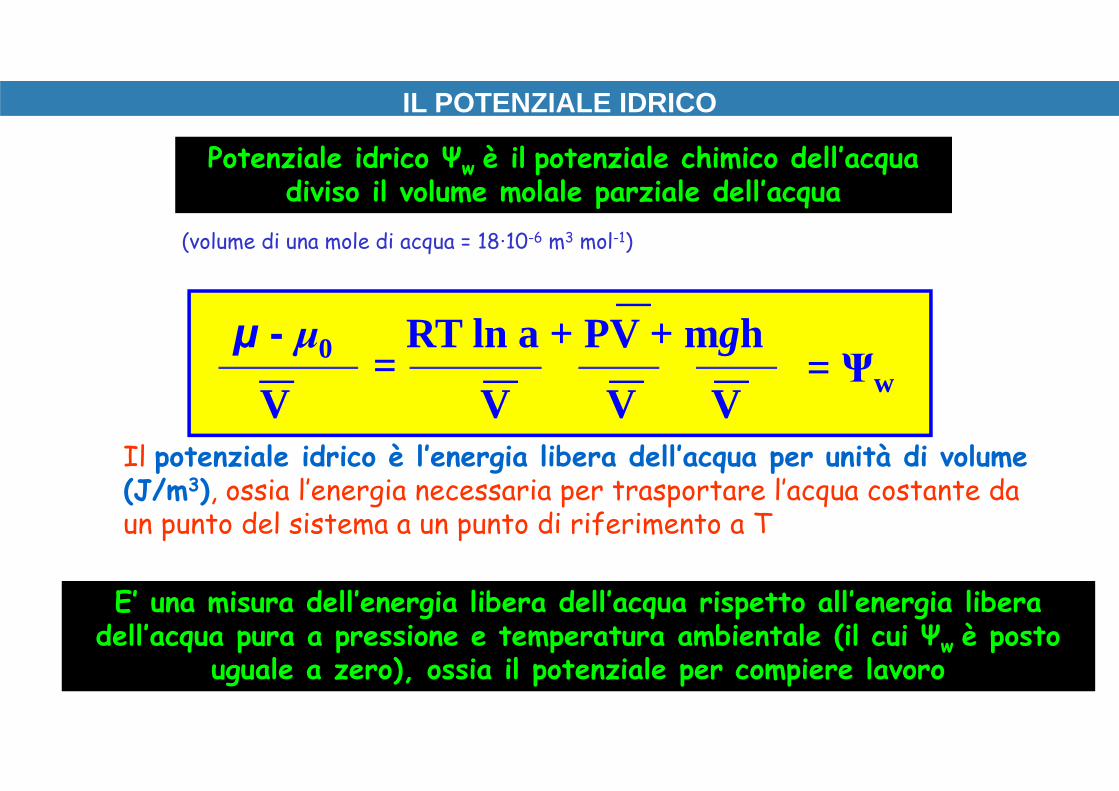
IL POTENZIALE IDRICO
μ - μ0 RT ln a + PV + mgh
V V V V= Ψw=
Potenziale idrico Ψw è il potenziale chimico dell’acqua diviso il volume molale parziale dell’acqua
Il potenziale idrico è l’energia libera dell’acqua per unità di volume (J/m3), ossia l’energia necessaria per trasportare l’acqua costante da un punto del sistema a un punto di riferimento a T
E’ una misura dell’energia libera dell’acqua rispetto all’energia libera dell’acqua pura a pressione e temperatura ambientale (il cui Ψw è posto
uguale a zero), ossia il potenziale per compiere lavoro
(volume di una mole di acqua = 18∙10-6 m3 mol-1)

Il potenziale idrico Ψw è tipicamente espresso in unità di pressione Pascal (Pa)che sono equivalenti alle unità J m-3
1 atm = 0,1013 MPa = 1,013 × 105 Pa
1 atm = 760 mm Hg
= 1.013 bar
H2O si muove sempre verso regioni a Ψw più basso ABBASSANDO LA PROPRIA ENERGIA LIBERA
All’equilibrio ΔΨw = 0
Ψiniziale > Ψfinale
Il potenziale idrico (ΨW) costituisce la forza motrice per il movimento dell’acqua nella pianta e definisce la direzione del
flusso idrico
IL POTENZIALE IDRICO

IL POTENZIALE DEL SOLUTO
RT ln aV
Ψs Potenziale del soluto o potenziale osmotico
Ψs rappresenta l’effetto su Ψw della presenza di soluti disciolti
�L’attività (frazione molare) dell’acqua pura è 1
�In una soluzione la frazione molare dell’acqua è < 1
Ψs è quindi sempre negativo

L’acqua tende a muoversi per diffusione da una soluzione a bassa concentrazione di soluti
(acqua ad alta concentrazione)ad una soluzione ad alta concentrazione di soluti
(acqua a bassa concentrazione)
IL POTENZIALE DEL SOLUTO
La diffusione in sistemi non omogenei è il trasferimento netto di particelle da regioni a più alta
concentrazione verso quelle a più bassa concentrazione. E’ molto veloce nelle miscele gassose, più
lenta nei sistemi liquidi. La diffusione in fase liquida è alla base dell’osmosi, che si verifica quando
in una soluzione liquida alcuni componenti possono diffondere liberamente rispetto ad altri che
non possono farlo.

IL POTENZIALE DEL SOLUTO
� I soluti aumentano l’entropia dell’acqua e ne diminuiscono l’energia libera rispetto a quella dell’acqua allo stato puro
� Poiché diminuiscono l’energia libera dell’acqua, i soluti riducono la capacità dell’acqua di compiere un lavoro
� I soluti diminuiscono il Potenziale Idrico
� Il Potenziale Osmotico è indipendente dalla natura del soluto

Equazione di van’t Hoff e Pressione Osmotica
Il potenziale osmotico (Ψs) si può esprimere in funzione della frazione molare dell’acqua o della concentrazione di soluto utilizzando l’equazione di
van’t Hoff
L’equazione di van’t Hoff permette di correlare il potenziale idrico alla pressione osmotica (π), pari al potenziale di soluto cambiato di segno
π = RTCs Ψs = - RTCs
π = RTCs = - RT ln aw = - Ψs
V

In acqua pura π = 0 Ψs = 0
In una soluzione π > 0 Ψs < 0
R: costante dei gas
T: temperatura assoluta
Cs: concentrazione dei soluti mol/L
Ψs = - π = - RTCs
Il segno negativo indica che i soluti disciolti riducono il potenziale idrico della soluzione
Equazione di van’t Hoff e Pressione Osmotica

POTENZIALE OSMOTICO NELL’ACQUA DI MARE
Fisiologia Vegetale – Taiz and Zeiger – Piccin Editore - Quarta Edizione Italiana

IL POTENZIALE DI PRESSIONE
P VV
= P ΨP Potenziale di pressione
Ψp si riferisce alla pressione idrostatica
Ψp si misura come differenza con la pressione atmosferica
P = Passoluta – Patmosferica
Quindi per l’acqua a pressione ambientale Ψp = 0 (anche se pressione assoluta = pressione atmosferica = 1 atm = 0.1 MPa)

IL POTENZIALE DI PRESSIONE
L’acqua tende a muoversi per flusso di massa da una zona a Pressione idrostatica maggiore verso una
zona a Pressione idrostatica minore
�Pressioni positive innalzano il Potenziale idrico Ψw
�Pressioni negative riducono il Potenziale idrico Ψw

Ψp può essere positivo, uguale a zero o negativo
Ψp è negativo nello xilema, quando la pianta sta traspirando, nelle pareti cellulari e nel suolo dove si può sviluppare una Tensione (o pressione idrostatica negativa) ~-0.6/-0.8 MPa nello xilema di radice e fusto
Ψp è positivo nelle cellule dove è definito Pressione di Turgore (o pressione idrostatica positiva) ~0.5 – 1 MPa
IL POTENZIALE DI PRESSIONE

EFFETTO DELLA GRAVITA’
m g hV
= ρw g h Ψg
Ψg si riferisce all’effetto della gravità su Ψw
Ψg dipende dalla densità dell’acqua (ρw), dall’accelerazione di gravità (g), e dall’altezza (h) dell’acqua rispetto allo stato di riferimento
ρw g = 0.01 MPa m-1Un’altezza di 10 metri si
traduce in un cambiamento di Ψw di 0.1 MPa

Ψw dipende dalla concentrazione, dalla pressione e dalla gravità
μ - μ0 RT ln a + PV + mgh
V V V V= Ψw=
Ψw = Ψs + Ψp + Ψg
Ψw = -π + P + ρgh

IL POTENZIALE IDRICO
Ψw = Ψs + Ψp + Ψg
Ψw = Ψs + Ψp Ψw = P - π
Se h < 5 – 10 m
Trasporto a livello cellulareΨg

La velocita’ del flusso
Il movimento dell’acqua:� in risposta ad un gradiente di concentrazione Ψs l’acqua si
muove per diffusione� in risposta ad un gradiente di pressione idrostatica Ψp l’acqua
si muove per flusso di massa� in risposta ad un gradiente di Potenziale idrico Ψw (che
tiene conto sia del gradiente di concentrazione sia del gradiente di pressione) l’acqua si muove per osmosi
Ψw = Ψs + Ψp

PRESSIONE OSMOTICA E OSMOSI
membrana permeabile all’acqua
pressione idrostatica
Pressione osmotica = pressione idrostaticarichiesta per prevenire un flusso netto di H 2O
L’osmosi si verifica quando due
soluzioni a concentrazione diversa
vengono separate da una
membrana semipermeabile.
Questa membrana si lascia
attraversare dal solvente ma non
dal soluto, la cui diffusione quindi
non può avvenire liberamente.
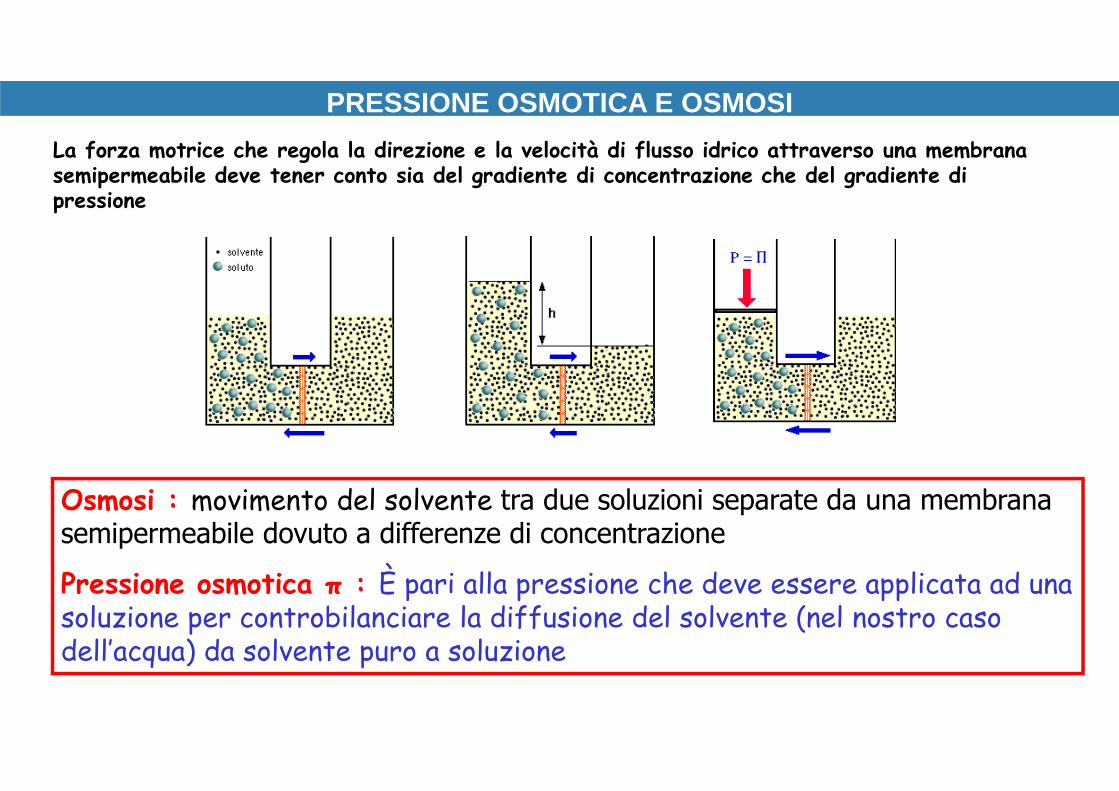
Osmosi : movimento del solvente tra due soluzioni separate da una membrana semipermeabile dovuto a differenze di concentrazione
Pressione osmotica π : È pari alla pressione che deve essere applicata ad una soluzione per controbilanciare la diffusione del solvente (nel nostro caso dell’acqua) da solvente puro a soluzione
PRESSIONE OSMOTICA E OSMOSI
La forza motrice che regola la direzione e la velocità di flusso idrico attraverso una membrana semipermeabile deve tener conto sia del gradiente di concentrazione che del gradiente di pressione

Mezzo Tipo di trasporto
Direzione
e Velocità
Forza guida
Velocità
Caratteristica del mezzo
Ambiente
libero
Diffusione molecolare
Gradiente di concentrazione
o di pressione di vapore
Coeff. Diffusione
Resistenza alla diffusione (rs + rb)
Flusso di massa
Gradiente di pressione
Viscosità
Raggio
Membrana semi-
permeabile
Osmosi Gradiente di potenziale idrico
Lp
(Conduttività idraulica della
membrana)
La velocita’ del flusso

3.7 Thermal motion of molecules leads to diffusion
Le molecole si rimescolano a seguito della loro agitazione termica casuale
Diffusione: Graduale mescolanza delle molecole e dissipazione finale delle differenze di concentrazione
Se il movimento per diffusione non avviene attraverso membrane semipermeabili, la forza guida è rappresentata dal gradiente di concentrazione dc/dx
Diffusione molecolare Alta concentrazione Bassa concentrazione

LA DIFFUSIONE
La velocità di trasporto dipende dalla forza motrice ΔCs/Δx e dalle caratteristiche fisiche del mezzo Ds
La prima legge di FICKJs = - Ds ΔCs
Δx
Js : velocità di trasporto del soluto (o densità di flusso) quantità di sostanza s che attraversa un’unità di area nell’unità di tempo
ΔCs/Δx gradiente di concentrazione
Ds : coefficiente di diffusioneè una costante di proporzionalità che misura la facilità di una sostanza s a muoversi attraverso un particolare mezzo
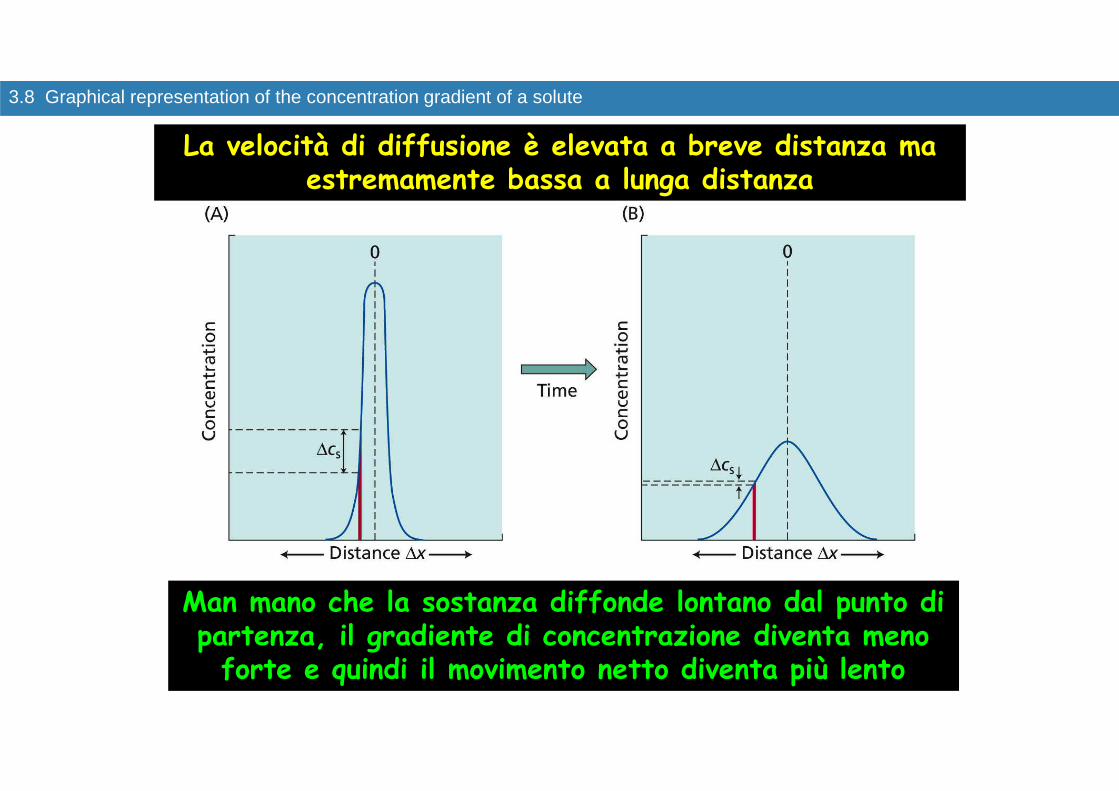
3.8 Graphical representation of the concentration gradient of a solute
Man mano che la sostanza diffonde lontano dal punto di partenza, il gradiente di concentrazione diventa meno forte e quindi il movimento netto diventa più lento
La velocità di diffusione è elevata a breve distanza ma estremamente bassa a lunga distanza

Il tempo impiegato da una sostanza per diffondere a una data distanza L dipende dal quadrato della distanza
Glucosio: Ds 10-9 m2 s-1 k = 1
L = 50 μm (dimensioni cellulari)
(Tc=1/2) = (50 10-6 m)2 / 10-9 m2 s-1 = 2,5 s
L = 1 m
(Tc=1/2) = (1m)2 / 10-9 m2 s-1 = 109 s = 32 anni
Il tempo impiegato dalla sostanza posta a qualsiasi distanza L dal punto di partenza per raggiungere la metà del valore di concentrazione del punto di partenza (t = ½) è data da:
(T c=1/2 )= L2 (k) /Ds

LA DIFFUSIONE
�La diffusione di piccole molecole in soluzione acquosa può essere efficace a dimensioni cellulari, ma è troppo lenta per il trasporto a lunga distanza
�La diffusione è di importanza fondamentale per la perdita del vapor d’acqua dalle foglie

FLUSSO DI MASSA
La velocità di trasporto dipende dalla forza motrice ΔP/Δx e dalle caratteristiche fisiche del mezzo πr4/8η
Gruppi di molecole si muovono insieme in risposta aun gradiente di pressionePressione alta -------> pressione minore
Spostamento dell’acqua all’interno delle tubazioni
Equazione di Poiseuille
Jv = (π r4 / 8η ) (ΔP/Δx)
[m3s-1]
Jv :velocità del flusso
r : raggio della tubatura
η : viscosità del liquidoPer H2O η = 10-9 MPa s-1

gradiente pressorio
ΔP = P1 – P2Jv = (π r4 / 8η ) (ΔP/Δx)
Pressione alta -------> pressione minore
Se il raggio raddoppia, la velocità del flusso del volume aumenterà di un fattore 16

IL MOVIMENTO DELL’ACQUA NELLE CELLULE VEGETALI
Per entrare o uscire da una cellula l’acqua deve attraversare
la membrana plasmatica, che ha le caratteristiche di una
membrana semipermeabile
NELLE CELLULE VEGETALI L’ACQUA SI MUOVE PER OSMOSI IN RISPOSTA
AL GRADIENTE DI POTENZIALE IDRICO Ψw
(CHE TIENE CONTO SIA DEL GRADIENTE DI CONCENTRAZIONE SIA DEL
GRADIENTE DI PRESSIONE)

IL POTENZIALE IDRICO DELLE CELLULE VEGETALI: LA PRE SSIONE DI TURGORE
� Il Potenziale del soluto è generato dalla presenza di soluti nei compartimenti cellulari, in particolare nel vacuolo, delimitati da membrane biologiche (semipermeabili)
� Il Potenziale di pressione è generato dalla presenza della parete cellulare, che esercita una pressione positiva sul protoplasto che prende il nome di pressione di turgore
� L’acqua si muove per diffusione attraverso la membrana plasmatica e per flusso di massa attraverso canali specifici chiamati acquaporine
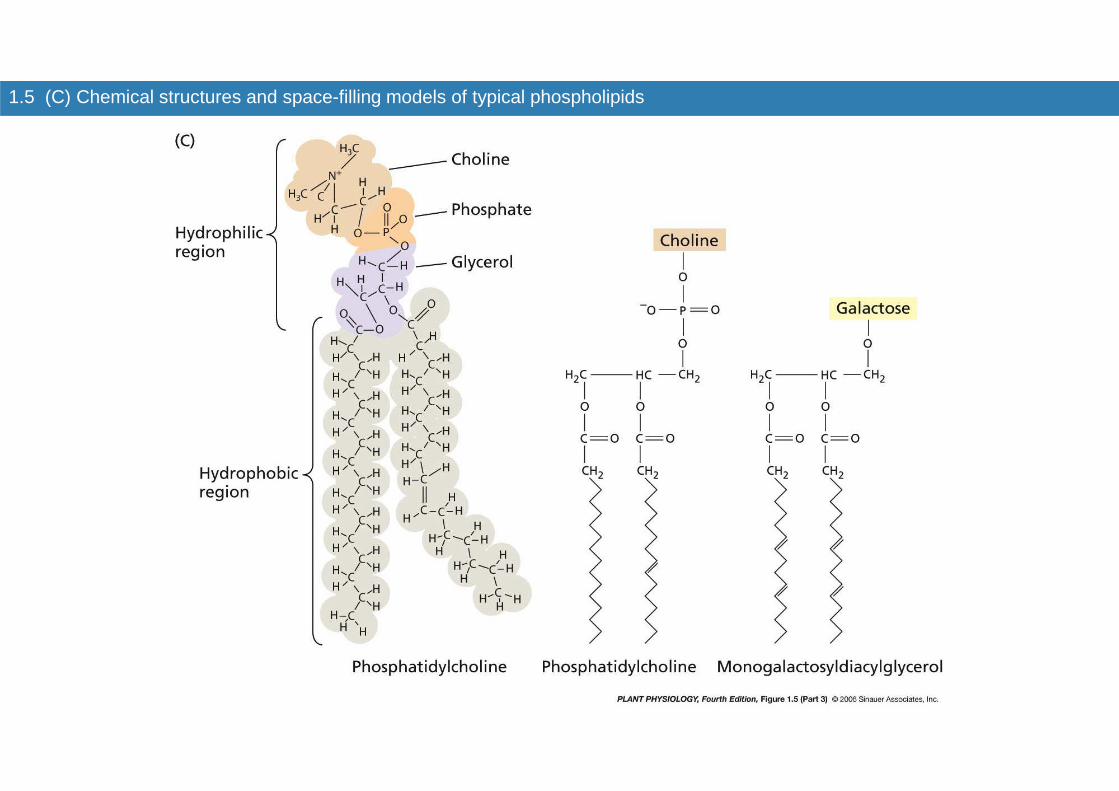
1.5 (C) Chemical structures and space-filling models of typical phospholipids

1.6 Different types of anchored membrane proteins

LE ACQUAPORINE
La semplice diffusione attraverso il bistrato lipidico non era sufficiente a giustificare la velocità di spostamento dell’acqua attraverso la membrana
Le acquaporine sono pori di membrana delineati da proteine Sono costituite da proteine integrali di membrana che formano canali selettivi per l’acqua
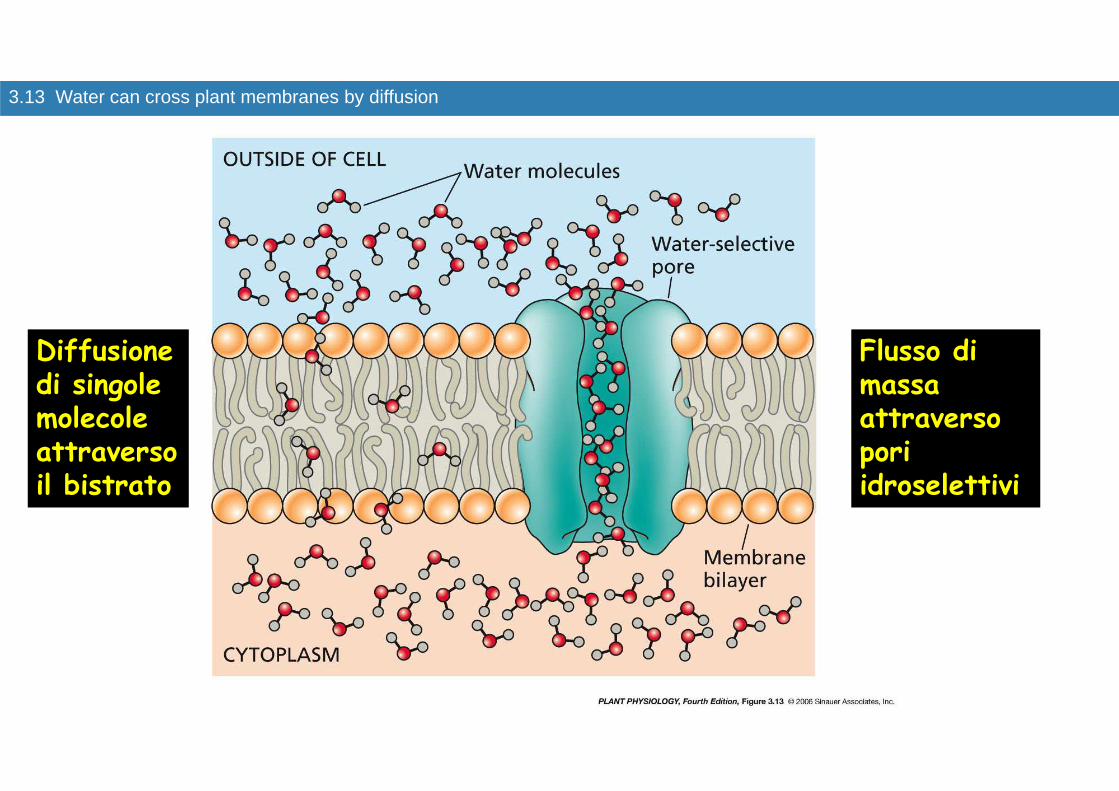
3.13 Water can cross plant membranes by diffusion
Diffusione di singole molecole attraverso il bistrato
Flusso di massa attraverso pori idroselettivi
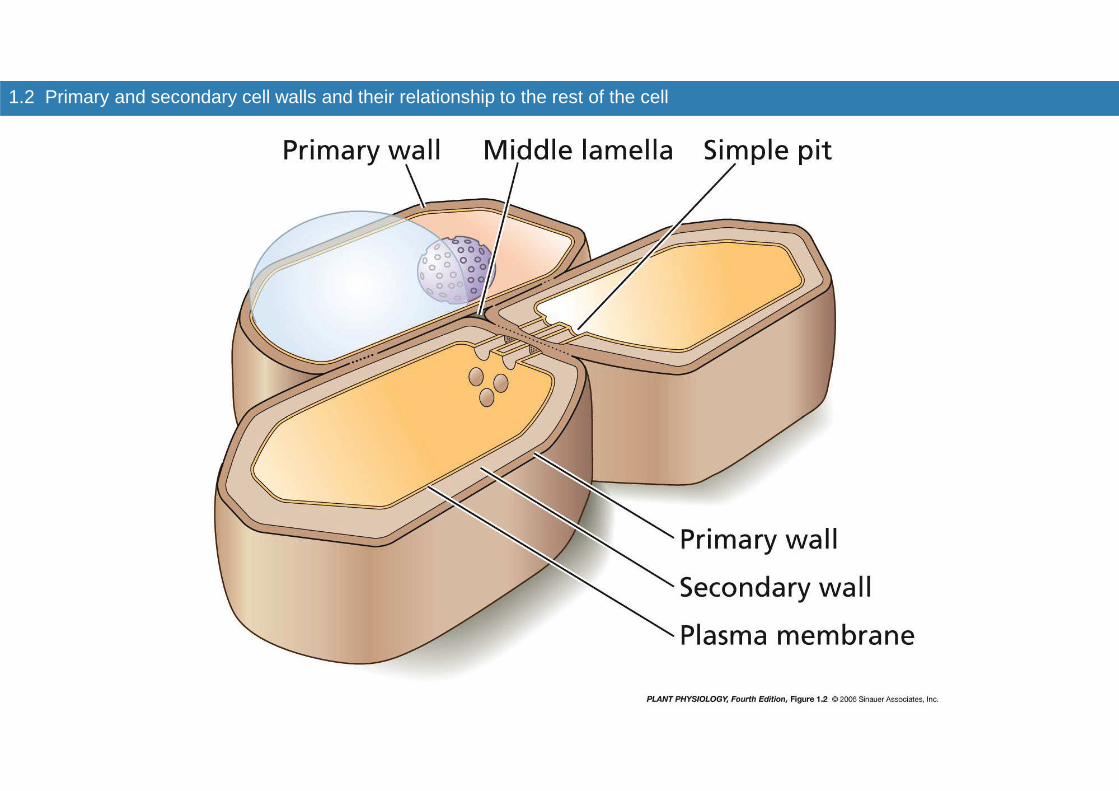
1.2 Primary and secondary cell walls and their relationship to the rest of the cell

CELLULE ANIMALI E CELLULE VEGETALI
Soluzione isotonicaSoluzione ipertonica: le cellule perdono acqua
Soluzione ipotonica : le cellule assumono acqua
Cellula animale (eritrocita)
Cellula vegetale
lisi
plasmolisi
la cellula raggrinzisce
Rigonfiamento e turgore
La membrana plasmatica si stacca dalla parete cellulare

Cellula turgida
La pressione di turgore crea una forza verso l’esterno, in modo uniforme e in tutte le direzioni
ESPANSIONE E PRESSIONE DI TURGORE NELLE CELLULE VEGE TALI
L’entità e la modalità dell’espansione cellulare dipendono da :
�Pressione di turgore
�Proprietà chimico-fisiche della parete, che può presentare zone di resistenza maggiore e zone di resistenza minore
velocità di assorbimento = ALpΔΨW

VARIAZIONI DI Ψw IN CELLULE ANIMALI E VEGETALI
� In cellule flaccide o prive di parete, come le cellule animali, i cambiamenti di Ψw sono dovuti principalmente a variazioni di Ψs
� In cellule turgide con pareti, come le cellule vegetali, i cambiamenti di Ψw sono dovuti principalmente a variazioni di Ψp

VELOCITA’ DI TRASPORTO DELL’ACQUA NELLA CELLULA
La velocità di trasporto (Jv) dipende dalla forza motrice (Gradiente di Ψw) e dalle caratteristiche fisiche del mezzo
(Conduttività idraulica della membrana Lp)
Velocità di flusso = forza motrice × conduttività idraulica
Jv = Lp (ΔΨw)
�Lp rappresenta la permeabilità della membrana
�Esprime con quanta facilità l’acqua può spostarsi
�È espressa in unità di volume per unità di area di membrana per unità di tempo per unità di forza motrice (m3
m-2 s-1 MPa-1)

3.12 The rate of water transport into a cell (Part 1)
Jv = Lp (ΔΨw)

3.12 The rate of water transport into a cell (Part 2)
Jv = Lp (ΔΨw)
A seguito dell’assorbimento d’acqua da parte della cellula, ΔΨw si abbassa causando il rallentamento della velocità di assorbimento di acqua con una cinetica di tipo esponenziale

LA VELOCITA’ DI ASSORBIMENTO DELL’ACQUA RALLENTA
t ½ =
0.693
(A) (Lp)
V
ε - Ψs
T ½ = tempo di dimezzamento
A = area della superficie cellulare
Lp = permeabilità della membrana
V = volume della cellula
ε = modulo elastico volumetrico
Ψs = potenziale osmotico cellulare
La velocità di flusso si dimezza in maniera
esponenziale al decrescere di ΔΨw, e il tempo
necessario affinché il tasso di velocità si
dimezzi è:

T ½ per il raggiungimento dell’equilibrio idrico ce llulare
Cellule con elevati rapporti superficie/volume, elevate conduttività di membrana e pareti cellulari rigide (ε elevato)
raggiungeranno rapidamente l’equilibrio con l’ambiente circostante
Il t1/2 della cellula varia tipicamente da 1 a 10 s
�Singole cellule raggiungono l’equilibrio del potenziale idrico con le cellule che le circondano in meno di un minuto
�Per tessuti multicellulari il t1/2 può essere superiore
Velocità di flusso del volume J = A Lp (ΔΨw) = L (ΔΨw)

Le proprietà chimico-fisiche della parete influiscono sulla perdita del turgore causata da riduzione del volume cellulare
Il modulo elastico volumetrico della cellula ε è una misura della rigidità della parete
�In cellule con pareti rigide (es. cellule del mesofillo) il valore di ε è elevato e i cambiamenti di volume associati alla perdita di turgore possono essere molto piccoli
�In cellule con pareti elastiche (es. cellule del parenchima acquifero) il valore di ε è basso e il cambiamento di volume associato alla perdita di turgore è sostanzialmente superiore
La pressione Ψp varia in funzione del volume della cellula
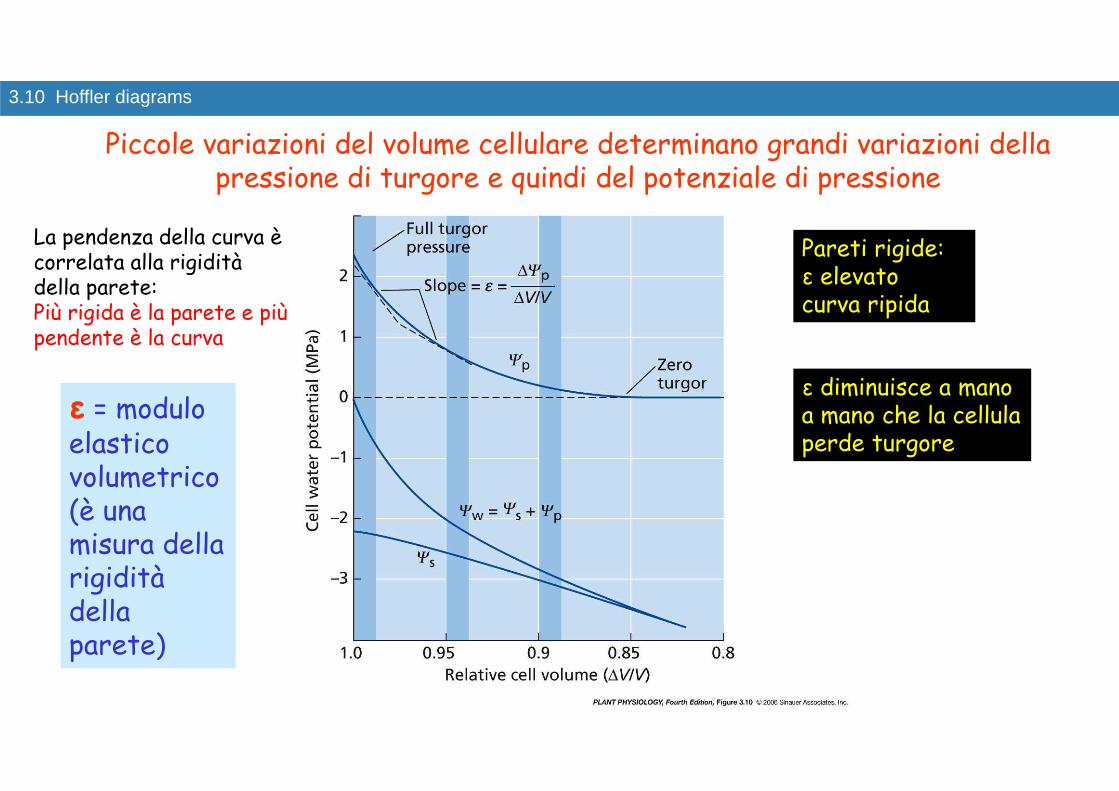
3.10 Hoffler diagrams
Pareti rigide: ε elevato curva ripida
ε diminuisce a mano a mano che la cellula perde turgore
ε = modulo elastico volumetrico(è una misura della rigidità della parete)
Piccole variazioni del volume cellulare determinano grandi variazioni della pressione di turgore e quindi del potenziale di pressione
La pendenza della curva è correlata alla rigidità della parete: Più rigida è la parete e più pendente è la curva

3.10 Hoffler diagrams
Piccole variazioni del volume cellulare determinano grandi variazioni della pressione di turgore e quindi del potenziale di pressione
Ψp si avvicina a 0 con perdite di volume del 10-15%
Se il volume cellulare diminuisce del 5%:
Ψp da 2.5 scende a 0.7 MPaΨs da -2.2 scende a -2.5 MpaΨw da >0 passa a -1.8 MPa

CURVA PRESSIONE/VOLUME
�Poiché le cellule vegetali posseggono pareti rigide, uncambiamento nel Ψw è di solito accompagnato da un grandecambiamento nel Ψp con piccoli cambiamenti nel volume delprotoplasto (perdita del 5-10% del volume cellulare Ψp = 0)
�Se la parete è molto rigida (grandi valori di ε) la curvapressione-volume risulta molto ripida e un piccolo cambiamento divolume porta ad un gran cambiamento nella pressione di turgorecellulare
�Per un cambiamento dell’1% del volume cellulare si ha unavariazione della pressione di turgore del 10%
�Quando ε e Ψp sono bassi (perdita del 10-15% del volumecellulare), i cambiamenti nel potenziale idrico sono dovutiprincipalmente alle variazioni di Ψs

3.11 Cross section of a cactus stem
ε (cellule dei tessuti che accumulano acqua) < ε (cellule fotosintetiche)
le cellule dei tessuti che accumulano acqua perdono un maggior contenuto d’acqua rispetto ad una cellula fotosintetica per ogni data diminuzione di Ψw
ε bassa: indica una buona elasticità della parete e il tessuto è adatto ad accumulare acqua
ε alta: il tessuto è rigido e non può accumulare acqua
Sezione trasversale del fusto di un cactus:Starto esterno fotosinteticoStrato interno non fotosintetico con funzione di accumulo di acqua.Durante la siccità l’acqua è persa dai tessuti non fotosintetici aiutando a mantenere lo status idrico dei tessuti fotosintetici

CELLULA VEGETALE E POTENZIALE IDRICO
In tutte le cellule vegetali vive ed idratate è presente una pressione intracellulare positiva, definita pressione di turgore (particolarmente elevata nei tubi cribrosi e
nelle cellule di guardia)
La pressione di turgore dipende:
�dalla natura semipermeabile della membrana plasmatica
� dalla natura elastica della parete cellulare primaria (che impedisce alle cellule di scoppiare e consente l’aumento della pressione idrostatica)

3.9 Five examples illustrating the concept of water potential and its components (Part 1)
Lo stato di riferimento utilizzato per definire il Ψw è l’acqua pura a pressione atmosferica e temperatura ambiente
In una soluzione 0.1 M di saccarosio il Ψw è
Ψs = -π = - RTCs

3.9 Five examples illustrating the concept of water potential and its components (Part 2)
Ψs = -πΨp = P
Ψw cell < Ψw solLa cellula assorbe acqua
All’equilibrio Ψw cell = Ψw sol Sviluppo di una pressione di turgore positiva

3.9 Five examples illustrating the concept of water potential and its components (Part 3)
Ψs = -πΨp = P
Ψw cell > Ψw solLa cellula perde acqua
All’equilibrio Ψw cell = Ψw solPlasmolisi (distacco del protoplasto dalla parete cellulare)

3.9 Five examples illustrating the concept of water potential and its components (Part 4)
Una forza esterna comprime la cellula senza alterare il potenziale idrico (osmosi inversa)
Si cambia il volume senza alterare il Ψw

















