

Il seme e il polline sono alcuni dei fattori di successo delle piante con semi.
Il seme (▶figura 26) è una struttura specializzata
costituita da un embrione racchiuso all’interno di un
rivestimento protettivo e corredato di una riserva
nutritiva che permetterà lo sviluppo della nuova
piantina. Nelle gimnosperme come nelle angiosperme, il
seme rappresenta dunque uno stadio di sopravvivenza
ben protetto. Alcune specie di piante producono semi
che possono restare vitali per molti anni (anche
migliaia) germinando soltanto in condizioni ambientali
propizie. Inoltre, molti semi hanno adattamenti
strutturali che favoriscono la disseminazione per mezzo
del vento o, più spesso, per mezzo di animali.
Figura 26. Il seme è un adattamento alla vita sulla terraferma.
(A) I semi del soffione, sono piccoli e leggeri, e possono
facilmente essere dispersi dal vento.
(B) Questi pinoli sono i semi di una pianta di pino: la parte
interna commestibile è la riserva nutritiva dell’embrione; il
rivestimento legnoso protegge l’embrione dalla disidratazione.
Questi semi sono adatti per resistere a lungo nel terreno.
Tra le piante con semi, solo i gruppi più antichi
possiedono gameti maschili natanti; tutte le altre
utilizzano il polline per trasferire sulle parti femminili
le cellule che daranno origine ai gameti maschili. Solo
dopo che il polline trasportato dal vento o dagli animali
ha raggiunto le strutture femminili (impollinazione) ha
luogo la fecondazione (▶figura 27).
Il punto di arrivo di questa straordinaria tendenza
evolutiva è costituito dalla completa indipendenza della
riproduzione rispetto all’acqua, che ha rappresentato un
enorme vantaggio evolutivo per la diffusione
nell’ambiente terrestre delle piante con semi.
Figura 27. I granuli pollinici sono i gametofiti maschili delle
piante a seme.
(A) Il polline di questa betulla si diffonde per mezzo del vento e
i granuli pollinici possono cadere in prossimità di un gametofito
femminile appartenente allo stesso albero o ad alberi diversi. (B)
Il polline di ogni specie differisce per forma e dimensione
(questa fotografia al microscopio elettronico è stata colorata
artificialmente).
Le piante con semi comprendono le gimnosperme e le angiosperme.
Le piante con semi, comparse nel tardo periodo
Devoniano circa 360 milioni di anni fa, derivarono
probabilmente da antiche felci legnose con semi e oggi
comprendono due grandi sottodivisioni: le
gimnosperme e le angiosperme.
Le gimnosperme (▶figura 28) sono le piante il cui seme
non è racchiuso in un frutto. Insieme alle felci e ad altre
piante senza semi, le gimnosperme formarono le grandi
foreste che hanno dominato il paesaggio per più di 200
milioni di anni durante il Mesozoico,
contemporaneamente ai dinosauri. Attualmente, il
gruppo più vasto delle gimnosperme è costituito
dalle conifere, che comprendono pini, abeti, sequoie e
molte altre specie con foglie aghiformi e strutture
contenenti i semi chiamate coni.
Le angiosperme presentano semi contenuti all’interno di
un frutto (▶figura 29). I semi delle angiosperme si
sviluppano a partire da strutture riproduttive molto
complesse, i fiori; per questo motivo esse sono
chiamate «piante con fiori». Le angiosperme
comparvero circa 140 milioni di anni fa e oggi
costituiscono la maggior parte delle piante moderne,
con 250000 specie sia arboree sia erbacee (▶figura 30).
5. Le Spermatofite sono piante vascolari dotate di semi.
Durante il Devoniano, quando erano già comparsi insetti, ragni, centopiedi e anfibi primitivi, alcune piante cominciarono a
proteggere i loro semi entro un involucro e a sviluppare fusti legnosi inspessiti, conquistando così grandi altezze e favorendo
la riproduzione. Insieme alla dominanza dello sporofito sul gametofito, queste due novità evolutive si rivelarono vincenti
nella conquista degli ambienti terrestri, oggi dominati proprio dalle piante con semi: le gimnosperme e le angiosperme.

Figura 28. Le conifere sono le gimnosperme più diffuse.
Nei climi più freddi le conifere formano estese foreste, come
questa in Siberia.
Figura 29. I semi delle angiosperme.
Diversamente da quelli delle gimnosperme, che sono «nudi», i
semi delle angiosperme sono «protetti» all’interno di un frutto
(come questo avocado).
Figura 30. La diversità delle angiosperme.
Le piante con fiore sono le più diffuse sul pianeta Terra, e
comprendono sia specie erbacee (A) sia specie arboree (B).
Le parole:
Gimnosperma deriva dal greco gymnós, «nudo»,
e spérma, «seme», per indicare che le piante non hanno
i semi protetti da una struttura specifica.
Angiosperma deriva dai termini greci angêion, «vaso», e
da spérma, «seme», sottolineando che i semi delle
piante con fiori sono custoditi in una struttura specifica,
il frutto, che ha origine dall’ovario.

Nelle gimnosperme, i coni producono gli ovuli e gli strobili producono il polline.
Le distese di abeti e cedri dell’America del Nord e quelle
di pini e larici dell’Eurasia sono tra le più vaste
formazioni forestali del mondo. Tutti questi alberi sono
conifere, cioè «portatrici di coni». Un cono (quello che
comunemente chiami pigna, (▶figura 31 A) è un
breve fusto lungo il quale sono inserite fittamente delle
squame legnose, ciascuna delle quali contiene
due ovuli; ogni ovulo è costituito da uno sporangio e da
un rivestimento protettivo.
Uno strobilo è un altro tipo di struttura coniforme più
piccola con squame derivate da foglie modificate
inserite lungo l’asse (▶figura 31 B). Ciascuna squama di
uno strobilo produce numerosi sporangi, ognuno dei
quali origina per meiosi molte spore aploidi; i gametofiti
maschili, o granuli pollinici, si sviluppano dalle spore.
Figura 31. I coni e gli strobili.
(A) Le scaglie dei coni femminili corrispondono a rami
modificati. (B) Le strutture contenenti le spore all’interno degli
strobili (ovvero i coni maschili) corrispondono a foglie
modificate.
Quando gli strobili sono maturi, le squame si aprono e
liberano una nuvola di polline costituita da milioni di
microscopici granuli (a primavera ti sarà capitato di
vedere i parabrezza delle automobili ricoperti da una
polvere giallastra: è il polline delle conifere). La parete
dei granuli pollinici contiene sporopollenina, il
composto biologico chimicamente più resistente che si
conosca, che ha il compito di proteggere il polline dalla
disidratazione e da eventuali danni chimici.
Il ciclo vitale delle gimnosperme.
Per illustrare il ciclo vitale delle gimnosperme ci
riferiamo a un pino (▶figura 32): l’albero è lo sporofito,
mentre il gametofito è ridotto a uno stadio
microscopico che si sviluppa all’interno dei coni.
I granuli di polline, trasportati dal vento, contengono le
cellule da cui si svilupperanno i gameti maschili.
Quando un granulo pollinico raggiunge l’ovulo su un
albero della stessa specie avviene l’impollinazione.
Dopo l’impollinazione, l’ovulo si divide per meiosi e
ciascuna spora aploide incomincia a svilupparsi in un
gametofito femminile. Occorrono mesi perché nei
gametofiti femminili si sviluppino le cellule uovo e nei
granuli di polline si sviluppino le cellule spermatiche.
A questo punto il granulo di polline produce un tubetto
pollinico che si allunga fino a raggiungere la cellula
uovo; le cellule spermatiche percorrono il tubetto, e,
una volta a contatto con la cellula uovo, innescano
la fecondazione.
Il processo dell’impollinazione e della fecondazione
richiede in tutto più di un anno. Dopo la fecondazione,
dallo zigote si sviluppa l’embrione dello sporofito e
l’intero ovulo si trasforma in seme. Il seme (▶figura 26
B) ha un rivestimento legnoso (il tegumento) e contiene
la riserva di cibo per l’embrione (costituita dai resti del
gametofito femminile). I coni dei pini liberano i semi
circa due anni dopo l’impollinazione; i semi cadono a
terra oppure vengono dispersi dal vento o dagli animali
e, quando le condizioni sono favorevoli, germinano (ciò
significa che gli embrioni incominciano a crescere). Alla
fine, dagli embrioni si sviluppano nuovi alberi.
6. Le gimnosperme sono le più antiche piante con semi.
La sottodivisione delle gimnosperme comprende le piante con semi che non producono fiori. Queste piante devono il nome
(che come abbiamo visto significa «a seme nudo») al fatto che i loro semi non sono protetti dal tessuto del frutto.
Attualmente esistono probabilmente meno di 850 specie di gimnosperme, in gran parte conifere; per quanto riguarda la loro
predominanza nell’ambiente terrestre, queste piante sono seconde soltanto alle angiosperme.

Le produzione dei gametofiti maschili sotto forma di
granuli di polline affranca completamente la pianta dalla
presenza di acqua per la riproduzione. L’ovulo è un
adattamento chiave che rappresenta uno strumento di
protezione per tutti gli stadi femminili del ciclo vitale; al
suo interno avvengono l’impollinazione, la
fecondazione e lo sviluppo dell’embrione. L’ovulo che si
trasforma in seme costituisce pertanto uno degli eventi
fondamentali a cui le gimnosperme e le angiosperme
devono il loro successo nella conquista della terraferma.
Nella maggior parte delle conifere l’unico tipo di
protezione dell’ovulo è fornito dalle squame, che sono
fittamente ammassate l’una contro l’altra e inserite
nell’asse del cono; alcune pigne hanno squame così
fitte e dure che soltanto il fuoco è in grado di aprirle e
indurre il rilascio dei semi.
Figura 32. Il ciclo vitale di una conifera.
Nelle conifere come il pino, ma anche in altre gimnosperme, i gametofiti hanno dimensioni microscopiche e dipendono completamente per il
nutrimento dallo sporofito. Nel seme di una gimnosperma sono presenti tessuti di 3 individui appartenenti a 3 diverse generazioni.

Alcune specie di conifere possiedono tessuti morbidi e
carnosi, simili a frutti, che circondano i semi, come le
«bacche» del tasso e del ginepro (▶figura 33); gli
animali possono cibarsi di questi tessuti e quindi
disperdere i semi attraverso le feci, spesso a grandi
distanze dalle piante di origine. Ma le «bacche» del
ginepro e del tasso non sono frutti veri e propri, che
invece sono strutture caratteristiche delle angiosperme.
Si tratta di un caso di analogia nel quale alcune
gimnosperme hanno evoluto una struttura che
superficialmente assomiglia a un frutto e svolge la
stessa funzione, ma che, studiata con attenzione, rivela
un’origine del tutto differente e indipendente.
Figura 33. Alcune gimnosperme possiedono strutture simili a
frutti.
Le bacche del ginepro rivestono i semi e servono a favorire la
loro dispersione, attirando gli animali che se ne nutrono. Queste
«pseudobacche», che si formano a partire da quattro squame
dello strobilo, non sono tuttavia veri frutti.
La presenza di tronchi legnosi ha permesso alle piante con semi di svilupparsi molto in altezza.
Le più antiche piante con semi producevano già il legno,
cioè unoxilema inspessito (▶figura 34), che permetteva
il sostegno per uno sviluppo in altezza maggiore
rispetto a quello di altre piante. La porzione più recente
del legno è adibita al trasporto di acqua, mentre il legno
più vecchio si irrobustisce con resina e altri materiali
per il sostegno, anche se non è più funzionale per la
conduzione.
Tuttavia, non tutte le piante con semi
sono legnose (cioè dotate di xilema): nel corso
dell’evoluzione molte di esse, dette erbacee, hanno
«perso» questo genere di accrescimento e quindi non
possono crescere molto in altezza, ma nonostante
questo sono riuscite a conquistare una vastissima
gamma di ambienti.
Figura 34. Gli anelli del legno.
Quando un albero viene tagliato, osservando la sezione del
tronco si notano degli anelli concentrici, che rappresentano
degli indicatori dell’età della pianta: ogni anello corrisponde a
un anno di vita.
Le gimnosperme attuali sono classificate in quattro classi: cicadine, ginkgofite, gnetofite e coniferofite.
Nonostante si conoscano poco meno di 850 specie
viventi, ancora oggi il gruppo delle gimnosperme
domina in alcune fasce climatiche terrestri, secondo
solo alle angiosperme. I rappresentanti delle quattro
classi odierne di gimnosperme non si somigliano molto
nell’aspetto. Esse sono:
Le cicadine (Cycadophyta), di cui sono note 140 specie,
sono piante tropicali e subtropicali simili a palme, alte
fino 20 m (▶figura 35); probabilmente si tratta del
gruppo più antico tra le gimnosperme esistenti. I loro
tessuti sono spesso fortemente tossici per l’uomo.
Figura 35. Le cicadine.
Molte cicadine possiedono una forma simile a quella delle felci e
delle palme, sebbene non siano strettamente imparentate con
nessuno di questi due gruppi.

Le ginkgofite (Ginkgophyta), assai comuni nell’era
Mesozoica, sono oggi rappresentate da un
unico genere con una sola specie, Ginkgo biloba, detta
anche albero dei ventagli (▶figura 36). Questa specie è
a sessi separati, con alberi maschili e alberi femminili.
Figura 36. Le ginkgofite.
Il caratteristico aspetto della pianta di Ginkgo biloba, delle sue
larghe foglie bilobate e il rivestimento carnoso del seme.
Le gnetofite (Gnetophyta) comprendono circa 90 specie
raccolte in tre generi molto diversi tra loro e che
condividono alcune caratteristiche con le
angiosperme. Welwitschia (▶figura 37), una gnetofita, è
una longeva pianta del deserto con foglie sfilacciate che
si allungano sulla sabbia raggiungendo anche i 3 m.
Figura 37. Le gnetofite.
Una Welwitschia mirabilis che cresce nel deserto della Namibia,
in Africa; queste piante possono vivere fino a 2000 anni. Le
foglie nastriformi si accrescono per tutta la vita e le loro
estremità si lacerano e si rompono progressivamente.
Le conifere (Coniferophyta) sono di gran lunga le
gimnosperme più abbondanti e diffuse, con circa 600
specie di piante che comprendono pini, abeti, larici,
cedri, cipressi, ginepri, tassi e sequoie (▶figura 38).
Esse sono tra le piante più grandi e longeve del pianeta
e formano foreste che coprono le regioni settentrionali e le
aree montane del pianeta e sono quindi fra le più imponenti
formazioni vegetali. Il loro nome significa portatrici di coni
(o strobili, cioè strutture assili recanti sporofilli). Tutte le
conifere sono eterosporee e gli sporofilli si trovano in coni
maschili e femminili separati e morfologicamente diversi,
che si formano all'apice dei rami. La maggior parte sono
piante sempreverdi, dotate di foglie aghiformi o ridotte a
piccole squame.
Figura 38. La diversità tra le gimnosperme.
Le conifere, come il pino (in alto) o il cedro in basso), sono le
piante dominanti in molte foreste attuali.
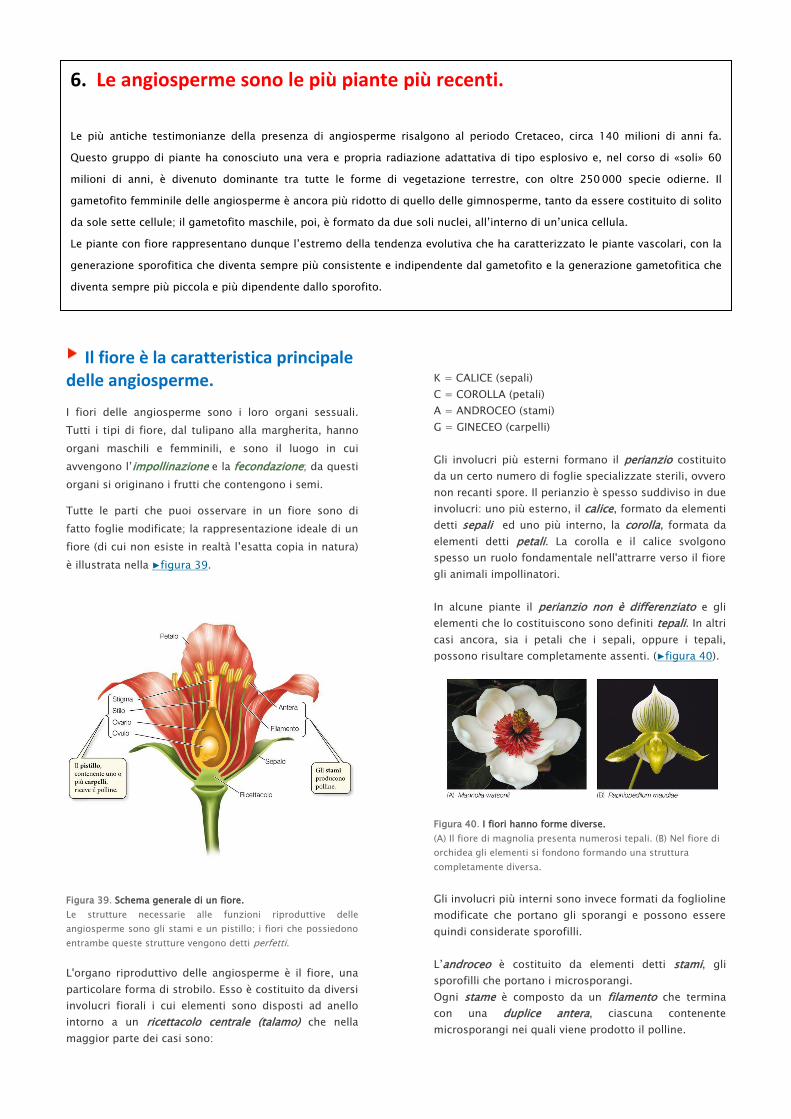
Il fiore è la caratteristica principale delle angiosperme.
I fiori delle angiosperme sono i loro organi sessuali.
Tutti i tipi di fiore, dal tulipano alla margherita, hanno
organi maschili e femminili, e sono il luogo in cui
avvengono l’impollinazione e la fecondazione; da questi
organi si originano i frutti che contengono i semi.
Tutte le parti che puoi osservare in un fiore sono di
fatto foglie modificate; la rappresentazione ideale di un
fiore (di cui non esiste in realtà l’esatta copia in natura)
è illustrata nella ▶figura 39.
Figura 39. Schema generale di un fiore.
Le strutture necessarie alle funzioni riproduttive delle
angiosperme sono gli stami e un pistillo; i fiori che possiedono
entrambe queste strutture vengono detti perfetti.
L'organo riproduttivo delle angiosperme è il fiore, una
particolare forma di strobilo. Esso è costituito da diversi
involucri fiorali i cui elementi sono disposti ad anello
intorno a un ricettacolo centrale (talamo) che nella
maggior parte dei casi sono:
K = CALICE (sepali)
C = COROLLA (petali)
A = ANDROCEO (stami)
G = GINECEO (carpelli)
Gli involucri più esterni formano il perianzio costituito
da un certo numero di foglie specializzate sterili, ovvero
non recanti spore. Il perianzio è spesso suddiviso in due
involucri: uno più esterno, il calice, formato da elementi
detti sepali ed uno più interno, la corolla, formata da
elementi detti petali. La corolla e il calice svolgono
spesso un ruolo fondamentale nell'attrarre verso il fiore
gli animali impollinatori.
In alcune piante il perianzio non è differenziato e gli
elementi che lo costituiscono sono definiti tepali. In altri
casi ancora, sia i petali che i sepali, oppure i tepali,
possono risultare completamente assenti. (▶figura 40).
Figura 40. I fiori hanno forme diverse.
(A) Il fiore di magnolia presenta numerosi tepali. (B) Nel fiore di
orchidea gli elementi si fondono formando una struttura
completamente diversa.
Gli involucri più interni sono invece formati da foglioline
modificate che portano gli sporangi e possono essere
quindi considerate sporofilli.
L’androceo è costituito da elementi detti stami, gli
sporofilli che portano i microsporangi.
Ogni stame è composto da un filamento che termina
con una duplice antera, ciascuna contenente
microsporangi nei quali viene prodotto il polline.
6. Le angiosperme sono le più piante più recenti.
Le più antiche testimonianze della presenza di angiosperme risalgono al periodo Cretaceo, circa 140 milioni di anni fa.
Questo gruppo di piante ha conosciuto una vera e propria radiazione adattativa di tipo esplosivo e, nel corso di «soli» 60
milioni di anni, è divenuto dominante tra tutte le forme di vegetazione terrestre, con oltre 250 000 specie odierne. Il
gametofito femminile delle angiosperme è ancora più ridotto di quello delle gimnosperme, tanto da essere costituito di solito
da sole sette cellule; il gametofito maschile, poi, è formato da due soli nuclei, all’interno di un’unica cellula.
Le piante con fiore rappresentano dunque l’estremo della tendenza evolutiva che ha caratterizzato le piante vascolari, con la
generazione sporofitica che diventa sempre più consistente e indipendente dal gametofito e la generazione gametofitica che
diventa sempre più piccola e più dipendente dallo sporofito.

Il gineceo è costituito da elementi detti carpelli, gli
sporofilli che recano i megasporangi. Dalla
trasformazione di uno o più carpelli si forma invece il
pistillo, nella cui parte basale, definita ovario, si trovano
gli ovuli. Nel pistillo si può inoltre riconoscere una parte
più allungata o stilo, la cui superficie distale, destinata a
ricevere il polline, prende il nome di stigma.
Un fiore che possiede tutti gli involucri fiorali è detto
completo, mentre uno che non li possiede tutti è detto
incompleto. Se un fiore produce sia megasporangi che
microsporangi, viene definito perfetto, viceversa, un
fiore che produce sporangi di un solo tipo è detto
imperfetto.
Molte angiosperme producono infatti due diversi tipi di
fiore, uno contenente soltanto megasporangi e l'altro
microsporangi. In questi fiori, di conseguenza, gli stami
o i carpelli non sono funzionanti, oppure mancano del
tutto.
Le specie in cui sono presenti sulla stessa pianta sia
fiori maschili che femminili vengono definite monoiche,
mentre quelle in cui i fiori maschili e femminili si
formano su piante diverse vengono dette dioiche; in
quest'ultimo caso esistono cioè piante femminili e
piante maschili.
Ogni specie è caratterizzata da una formula fiorale, la
quale indica quali sono gli involucri che costituiscono il
fiore e quanti sono gli elementi che li costituiscono.
Ad esempio, la formula fiorale K5C5A10G1, indica che il
fiore di una data specie è formato da un calice che
possiede 5 sepali, una corolla che possiede 5 petali, un
androceo che possiede 10 stami e un gineceo che
possiede un solo carpello.
Per quanto riguarda la disposizione, i fiori si possono
presentare singolarmente oppure essere raggruppati a
formare un’infiorescenza, per esempio l’ombrella delle
ombrellifere (la famiglia che comprende la carota e il
prezzemolo), i capolini delle asteracee (la famiglia
delle margherite e del girasole) e le spighe di molte
graminacee (▶figura 41).
Figura 41. Le infiorescenze.
(A) L’infiorescenza del dauco (un membro della famiglia delle
ombrellifere) è un’ombrella composta, formata da fiori sorretti
da steli che si originano in un punto centrale comune. (B) I
girasoli appartengono alla famiglia delle asteracee (o
composite). Nel capolino, ciascuna struttura allungata, simile a
un petalo, è in realtà un fiore, mentre la porzione centrale è
formata da dozzine o centinaia di altri fiori. (C) Le piante
erbacee come queste formano infiorescenze definite spighe.
Evoluzione del fiore.
I botanici manifestano tuttora opinioni contrastanti su
quale sia il tipo di fiore evolutivamente più primitivo. È
comunque ormai certo che i primi fiori possedevano
numerosi tepali (o sepali e petali), carpelli e stami, tutti
disposti a spirale intorno alla parte centrale.
Nel corso dell'evoluzione, questa struttura di base si è
poi modificata e a seconda dei casi si è verificata una
differenziazione dei petali dai sepali, la formazione di
un numero fisso di organi, la loro disposizione ad
anello o, infine, il passaggio da una simmetria radiale
(come quella dei gigli) in cui esistono molti piani di
simmetria che dividono il fiore in due parti equivalenti,
a una simmetria bilaterale (come quella dei fiori del
pisello o delle orchidee) in cui esiste un solo piano di
simmetria che divide il fiore in due parti che sono l’una
l’immagine speculare dell’altra. In molti casi la
simmetria bilaterale è accompagnata spesso da estese
fusioni fra le varie parti.
I primi carpelli rappresentavano sicuramente foglie
modificate e apparivano come sporofilli piegati ma non
completamente chiusi, di una forma intermedia tra
quella tipica delle gimnosperme e quella delle attuali
angiosperme. Successivamente, i carpelli si sono fusi e
sono stati progressivamente inglobati all'interno del
ricettacolo. Nei fiori delle angiosperme, evolutisi più
recentemente, le altre parti fiorali risultano disposte
all'estremo apicale dell'ovario piuttosto che alla base.
Anche gli stami dei primitivi fiori dovevano possedere
una struttura a forma di foglia, ed essere quindi
piuttosto diversi da quelli schematizzati in un ipotetico
fiore. In molti fiori attuali il pistillo presenta uno stilo di
forma allungata e anche le antere sono situate all'apice
di lunghi filamenti. L'allungamento di queste strutture è
stato probabilmente favorito dalla selezione naturale,
poiché esso aumenta la probabilità di impollinazione. La
presenza di lunghi peduncoli rende infatti più agevole il
contatto delle antere con il corpo degli insetti
impollinatori o, nel caso di impollinazione anemofila,
espone più efficacemente tali strutture all'azione del
vento. Argomentazioni simili valgono con ogni
probabilità anche per spiegare lo sviluppo di uno stilo di
forma allungata.
Recentemente, tuttavia, è stata proposta una teoria
alternativa, che interpreta l'allungamento dello stilo in
termini di selezione sessuale. Secondo quest'ipotesi,
l'allungamento dello stilo permetterebbe alla pianta
femminile di selezionare quei granuli pollinici in cui si è
realizzato un maggiore allungamento del tubetto
pollinico, e ciò come indizio di un patrimonio genetico
maschile con caratteristiche complessivamente
«migliori».

Le parole:
Petalo deriva dal greco petannýnai, «aprire»,
mentre sepalo è stato coniato sullo stesso modello,
derivandolo dal latino saepes, «recinto». Il senso è che i
sepali fanno da recinzione all’ingresso nel fiore,
segnato dai petali.
Monóico e dióico derivano dal greco oikós, «casa» (da
cui derivano anche ecologia ed economia), con i
suffissi mono- e di-, nel senso di fiori che ospitano uno
o due apparati sessuali.
Carpello deriva dal termine greco karpós, «frutto», e
infatti indica una o più foglie modificate che
proteggono il frutto.
Le angiosperme si sono coevolute con i loro impollinatori.
Mentre molte gimnosperme sono impollinate grazie al
vento (impollinazione anemofila), che trasporta
«nuvole» di granuli di polline, la maggior parte
delle angiosperme è impollinata da animali
(impollinazione zoofila). Moltissimi fiori favoriscono la
visita degli impollinatori offrendo loro del cibo,
il nettare zuccherino (ma anche i granuli di polline
stessi possono essere ricercati come nutrimento dagli
animali), spesso attirandoli con un particolare profumo.
Durante la visita, gli animali si imbrattano di polline,
che trasportano di fiore in fiore e di pianta in pianta,
favorendo in modo attivo l’aumento della varietà
genetica delle popolazioni vegetali. Gli animali
impollinatori più importanti sono senza dubbio gli
insetti, in particolare le api (▶figura 42 A); anche alcuni
uccelli, come i colibrì (▶figura 42 B) e certi pipistrelli
svolgono questo ruolo benefico.
Figura 42. L’impollinazione zoofila.
Molte specie di angiosperme si affidano agli animali per la
dispersione del polline.
Nell’ambiente terrestre le angiosperme e i loro
impollinatori si sono evoluti plasmandosi a vicenda per
oltre 130 milioni di anni, un processo
chiamato coevoluzione: gli animali hanno influenzato
l’evoluzione delle piante e le piante hanno influenzato
l’evoluzione dei loro impollinatori. Solo così, con questo
tipo di pressione adattativa reciproca, si può spiegare
l’incredibile diversità e specificità che si osserva tra fiori
e animali impollinatori. Per esempio, alcune specie
di Yucca (il genere cui appartiene il tronchetto della
felicità, comune pianta d’appartamento) in natura sono
impollinate da un’unica specie di falena (una farfalla
notturna). Un’impollinazione di questo tipo, affidata a
poche specie o addirittura soltanto a una specie
animale, fornisce alla pianta un meccanismo perfetto
per il trasferimento di polline da un individuo all’altro
(anche se la rende vulnerabile in caso di diminuzione o
scomparsa dei suoi impollinatori specialisti).
La maggior parte delle interazioni pianta-impollinatore
è tuttavia molto meno specifica: infatti la stessa specie
di angiosperma viene di solito impollinata da molte
specie animali, e lo stesso impollinatore funge da
veicolo di polline per molte piante diverse.
I fiori impollinati dagli uccelli sono spesso di colore
rosso e privi di odore, mentre quelli visitati da insetti
hanno odori caratteristici; addirittura, molti fiori
impollinati dalle api hanno disegni peculiari sui petali,
segnati da vere e proprie «piste di atterraggio», le
cosiddette «guide del nettare», che si rendono evidenti
solo nella regione ultravioletta dello spettro della luce,
proprio quella in cui le api vedono meglio.
Le caratteristiche del ciclo vitale delle angiosperme.
Come abbiamo appena visto, le principali caratteristiche
delle angiosperme si possono sintetizzare in cinque
punti principali:
1. doppia fecondazione;
2. produzione di endosperma;
3. ovuli e semi racchiusi in un carpello;
4. presenza di fiori;
5. presenza di frutti.
Nelle angiosperme, l’impollinazione consiste nell’arrivo
di un granulo di polline all’interno di un fiore; questo è
soltanto il primo dei tre eventi che porteranno alla
formazione di un seme. Seguono quindi la formazione
del tubetto pollinico (▶figura 43) la successiva
fecondazione, che ha la peculiarità di essere «doppia».
Figura 43. I tubetti pollinici iniziano a svilupparsi.
Questi granuli pollinici hanno raggiunto strutture a forma di
dito presenti sullo stigma di un fiore di Arabidopsis, e i loro
tubetti pollinici sono penetrati al suo interno.
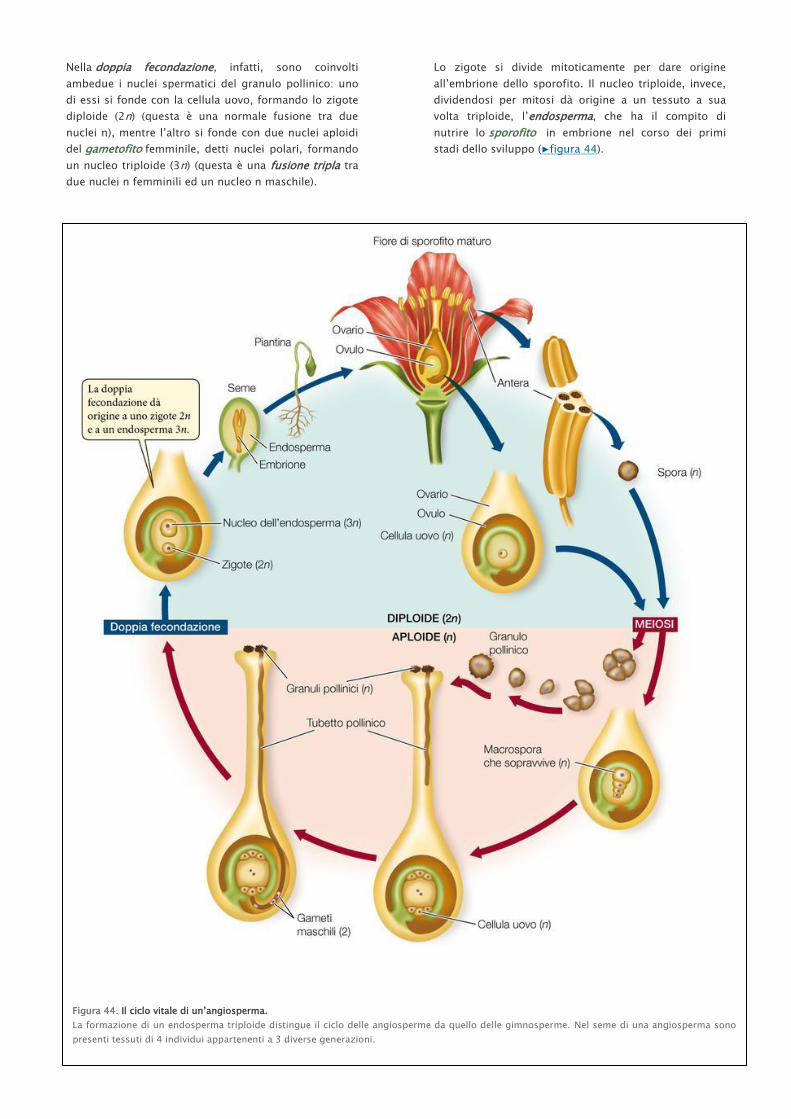
Nella doppia fecondazione, infatti, sono coinvolti
ambedue i nuclei spermatici del granulo pollinico: uno
di essi si fonde con la cellula uovo, formando lo zigote
diploide (2n) (questa è una normale fusione tra due
nuclei n), mentre l’altro si fonde con due nuclei aploidi
del gametofito femminile, detti nuclei polari, formando
un nucleo triploide (3n) (questa è una fusione tripla tra
due nuclei n femminili ed un nucleo n maschile).
Lo zigote si divide mitoticamente per dare origine
all’embrione dello sporofito. Il nucleo triploide, invece,
dividendosi per mitosi dà origine a un tessuto a sua
volta triploide, l’endosperma, che ha il compito di
nutrire lo sporofito in embrione nel corso dei primi
stadi dello sviluppo (▶figura 44).
Figura 44. Il ciclo vitale di un’angiosperma.
La formazione di un endosperma triploide distingue il ciclo delle angiosperme da quello delle gimnosperme. Nel seme di una angiosperma sono
presenti tessuti di 4 individui appartenenti a 3 diverse generazioni.

Le angiosperme producono frutti, che contengono i semi.
Dopo la fecondazione, l’ovario di un fiore insieme
ai semi che contiene è destinato a trasformarsi in
un frutto. Questa struttura non soltanto protegge i semi
ma può anche favorirne la dispersione attirando animali
che se ne nutrono o su cui resta impigliata. Un frutto
può derivare soltanto dall’ovario maturo insieme ai suoi
semi oppure comprendere altre parti del fiore o
strutture associate ad esso.
Un frutto semplice, come per esempio una ciliegia
(▶figura 45 A), deriva da un carpello singolo o da
diversi carpelli riuniti; il lampone è invece un esempio
di frutto aggregato, o composto (▶figura 45 B), che si
sviluppa cioè da numerosi carpelli separati di un singolo
fiore; gli ananas e i fichi sono esempi di frutti
multipli (▶figura 45 C), formati a partire da
un’infiorescenza. I frutti derivati da parti diverse dal
carpello e dai semi sono chiamati falsi frutti (▶figura 45
D); sono esempi di questo tipo di frutto le mele, le pere
e le fragole.
Il processo mediante il quale i semi vengono dispersi,
fino a raggiungere un ambiente adatto alla
germinazione, è chiamato disseminazione.
In molte piante questo significa semplicemente la
caduta per gravità del frutto e la sua successiva
disgregazione, ma in altri casi può essere anche più
complessa. Come abbiamo già visto nel caso
dell’impollinazione, possono essere coinvolti agenti di
dispersione come il vento (chiamato
trasporto anemocoro), che porta il frutto lontano dalla
pianta madre grazie alle sue caratteristiche
aerodinamiche (vedi ▶figura 26 A).
Nel trasporto zoocoro invece vengono utilizzati gli
animali, come per esempio se i frutti sono spinosi e si
agganciano al pelo dei mammiferi, oppure se i frutti
sono carnosi e vengono ingeriti da animali che ne
disperdono i semi. Addirittura, i semi di alcune specie
per poter germinare devono prima entrare in contatto
con i succhi gastrici di un animale. Infine, quando il
mezzo di disseminazione è l’acqua si parla di
trasporto idrocoro; un esempio particolare è la noce di
cocco, che può galleggiare ed essere trasportata per
chilometri dalle correnti marine.
Le parole:
Frutto è un termine che in botanica indica tutto ciò che
deriva dai carpelli o, in senso più ampio, da una parte
del fiore. Pertanto sono frutti anche diversi ortaggi,
come i pomodori o i peperoni.
Figura 45. I frutti possiedono forme, colori, profumi e sapori diversi.
(A) La ciliega è un frutto semplice. (B) Il lampone è un frutto composto. (C) Un frutto multiplo, l’ananas, e (D) un falso frutto, la fragola.

Il successo evolutivo delle angiosperme.
Le angiosperme sono le piante più diffuse sulla Terra da
più di 100 milioni di anni. Attualmente se ne conoscono
circa 250 000 specie; la maggior parte del nostro cibo
deriva però dai prodotti di poche centinaia di specie: le
radici (carote e barbabietole), i frutti (mele, uva, noci,
lamponi e zucche), i semi delle leguminose (piselli e
fagioli), oppure i semi dei cereali come il riso, il grano e
il mais.
I botanici suddividono le angiosperme in due gruppi, le
monocotiledoni e le dicotiledoni (▶figura 46). I nomi
«monocotiledone» e «dicotiledone» si riferiscono alle
prime foglie che compaiono nell’embrione della pianta:
queste foglioline embrionali sono chiamate cotiledoni.
L’embrione delle monocotiledoni possiede una foglia
sola, mentre quello delle dicotiledoni ne ha due.
Le monocotiledoni comprendono circa 65 000 specie
tra cui le orchidee, i bambù, le palme, i gigli, ma anche i
cereali e altre piante erbacee (▶figura 47); il cotiledone
singolo è visibile all’interno del seme. Anche le foglie, i
fusti, i fiori e le radici delle monocotiledoni hanno delle
caratteristiche particolari: nella maggior parte delle
monocotiledoni, le foglie presentano nervature parallele
e i tessuti vascolari all’interno del fusto (che trasportano
l’acqua e le sostanze nutritive) sono organizzati in fasci
con una disposizione sparsa; i petali e gli altri elementi
del fiore sono tre o multipli di tre.
La maggior parte delle angiosperme è però costituita
da dicotiledoni. Sono dicotiledoni gran parte degli
arbusti e degli alberi, nonché la maggior parte delle
piante ornamentali e molte piante di uso alimentare
(▶figura 48). Le foglie delle dicotiledoni possiedono una
rete ramificata di nervature e gli steli presentano
dei fasci vascolari che, in sezione trasversale, sono
disposti a formare un anello periferico; i petali e le altre
parti del fiore di solito sono quattro o cinque, oppure
multipli di essi.
Figura 46. Monocotiledoni e dicotiledoni a confronto.
La presenza di un solo cotiledone è uno dei caratteri che distinguono le monocotiledoni dalle altre angiosperme.

Figura 47. Le monocotiledoni.
(A) Le palme sono fra le poche monocotiledoni arboree. (B) Molte piante erbacee, come il grano, sono monocotiledoni. (C) Le monocotiledoni
includono inoltre alcuni comuni fiori da giardino, come questi gigli.
Figura 48. Le dicotiledoni.
(A) La famiglia delle cactacee costituisce un grande gruppo di dicotiledoni, con circa 1500 specie soltanto nelle Americhe. (B) Le rose
rampicanti sono dicotiledoni che appartengono alla famiglia delle rosacee, così come le rose coltivate.

Gli organi delle angiosperme formano un sistema aereo e un sistema radicale
Le angiosperme possiedono tre tipi di
organi vegetativi (ossia non coinvolti nella
riproduzione): le radici, i fusti e le foglie. Tutti gli organi
sono organizzati a formare due apparati ben distinti:
il sistema aereo e il sistema radicale. I piani strutturali
fondamentali di una monocotiledone e di una
dicotiledone tipica sono illustrati nella ▶figura 49:
Il sistema aereo di una pianta è formato dai fusti,
dalle foglie e dai fiori. In linea generale,
le foglie costituiscono gli organi principali adibiti
alla fotosintesi. I fusti sostengono le foglie, le
espongono alla luce del Sole e forniscono le vie per
il trasporto dei materiali tra le radici e le foglie.
I nodi corrispondono ai punti di inserimento delle
foglie sul fusto, mentre le zone del fusto tra nodi
successivi vengono definite internodi.
Il sistema radicale àncora la pianta al terreno e
fornisce nutrimento al corpo vegetale. L’estrema
ramificazione delle radici e il loro elevato rapporto
superficie/volume permettono di assorbire l’acqua
e i nutrienti dal terreno.
Figura 49. Gli organi e gli apparati vegetativi.
La struttura corporea di base e i principali organi vegetativi
delle monocotiledoni e delle dicotiledoni.
Dal fusto hanno origine le gemme e i fiori.
La funzione principale del fusto è quella di portare in
alto gli organi riproduttivi (i fiori) e quelli fotosintetici
(foglie). A differenza delle radici, il fusto porta gemme
di vario tipo: una gemma è un germoglio allo stato
embrionale. In corrispondenza dei nodi si sviluppano le
foglie e dove la foglia si inserisce sul fusto si trova
una gemma ascellare (vedi ▶figura 49). Quando la
gemma si attiva, può svilupparsi in un nuovo ramo o in
un’estensione del sistema aereo.
All’apice di ogni fusto o ramo di trova una gemma
apicale, che produce le cellule responsabili
dell’accrescimento in altezza o in larghezza e dello
sviluppo del fusto.
Le angiosperme presentano sistemi conduttori articolati.
Un’altra caratteristica delle angiosperme è la presenza
di sistemi conduttori articolati: lo xilema infatti
possiede speciali cellule conduttrici
chiamate trachee o elementi dei vasi (▶figura 50 A), di
ampio diametro e connesse tra loro, che formano lunghi
condotti senza interruzioni; l’altra cellula tipica dello
xilema delle angiosperme è la fibra, che svolge un ruolo
fondamentale nel sostegno. Il floema delle piante con
fiori è costituito da particolari tipi di cellule
chiamate tubi cribrosi (▶figura 50 B).
Figura 50. La conduzione nelle angiosperme.
(A) Elementi tracheali nel fusto di una pianta di zucca: le pareti
sono colorate in rosso (si noti il diverso tipo di inspessimento
ad anello e spirale). (B) Elementi dei tubi cribrosi, adibiti alla
conduzione delle sostanze nutritive nel fusto di una pianta di
cetriolo. (C) Floema e xilema sono organizzati in fasci, come si
vede in questa sezione di fusto.
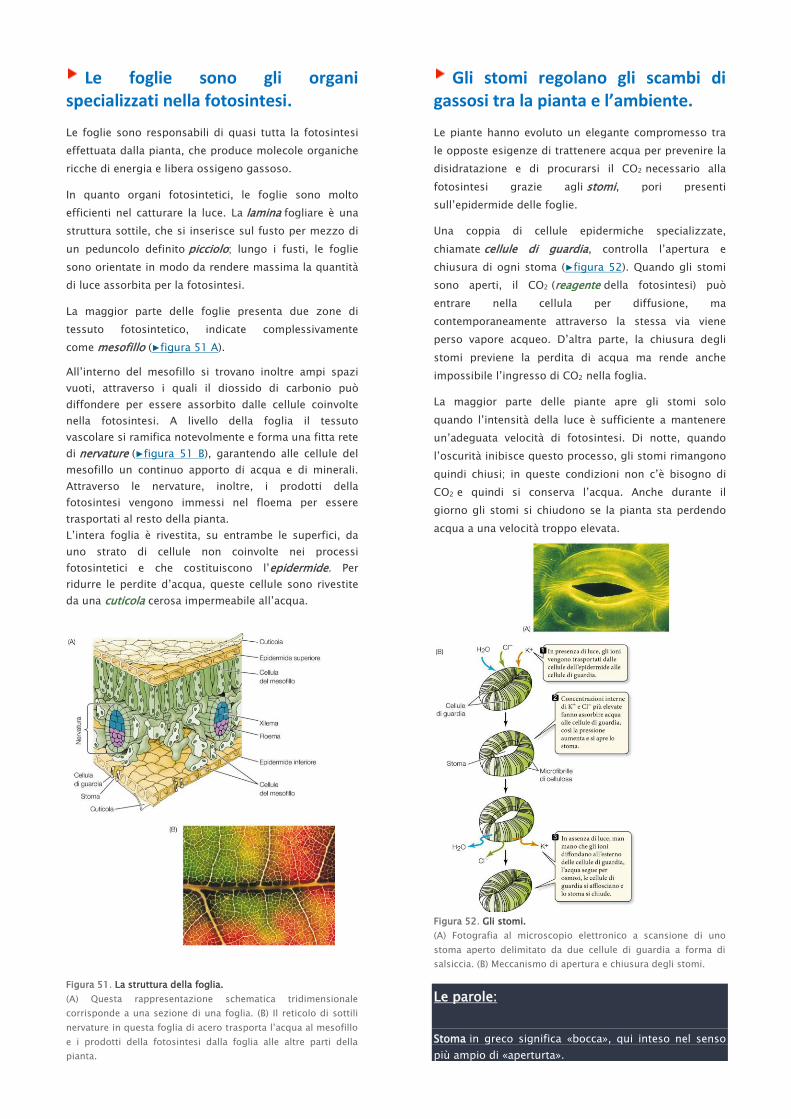
Le foglie sono gli organi specializzati nella fotosintesi.
Le foglie sono responsabili di quasi tutta la fotosintesi
effettuata dalla pianta, che produce molecole organiche
ricche di energia e libera ossigeno gassoso.
In quanto organi fotosintetici, le foglie sono molto
efficienti nel catturare la luce. La lamina fogliare è una
struttura sottile, che si inserisce sul fusto per mezzo di
un peduncolo definito picciolo; lungo i fusti, le foglie
sono orientate in modo da rendere massima la quantità
di luce assorbita per la fotosintesi.
La maggior parte delle foglie presenta due zone di
tessuto fotosintetico, indicate complessivamente
come mesofillo (▶figura 51 A).
All’interno del mesofillo si trovano inoltre ampi spazi
vuoti, attraverso i quali il diossido di carbonio può
diffondere per essere assorbito dalle cellule coinvolte
nella fotosintesi. A livello della foglia il tessuto
vascolare si ramifica notevolmente e forma una fitta rete
di nervature (▶figura 51 B), garantendo alle cellule del
mesofillo un continuo apporto di acqua e di minerali.
Attraverso le nervature, inoltre, i prodotti della
fotosintesi vengono immessi nel floema per essere
trasportati al resto della pianta.
L’intera foglia è rivestita, su entrambe le superfici, da
uno strato di cellule non coinvolte nei processi
fotosintetici e che costituiscono l’epidermide. Per
ridurre le perdite d’acqua, queste cellule sono rivestite
da una cuticola cerosa impermeabile all’acqua.
Figura 51. La struttura della foglia.
(A) Questa rappresentazione schematica tridimensionale
corrisponde a una sezione di una foglia. (B) Il reticolo di sottili
nervature in questa foglia di acero trasporta l’acqua al mesofillo
e i prodotti della fotosintesi dalla foglia alle altre parti della
pianta.
Gli stomi regolano gli scambi di gassosi tra la pianta e l’ambiente.
Le piante hanno evoluto un elegante compromesso tra
le opposte esigenze di trattenere acqua per prevenire la
disidratazione e di procurarsi il CO2 necessario alla
fotosintesi grazie agli stomi, pori presenti
sull’epidermide delle foglie.
Una coppia di cellule epidermiche specializzate,
chiamate cellule di guardia, controlla l’apertura e
chiusura di ogni stoma (▶figura 52). Quando gli stomi
sono aperti, il CO2 (reagente della fotosintesi) può
entrare nella cellula per diffusione, ma
contemporaneamente attraverso la stessa via viene
perso vapore acqueo. D’altra parte, la chiusura degli
stomi previene la perdita di acqua ma rende anche
impossibile l’ingresso di CO2 nella foglia.
La maggior parte delle piante apre gli stomi solo
quando l’intensità della luce è sufficiente a mantenere
un’adeguata velocità di fotosintesi. Di notte, quando
l’oscurità inibisce questo processo, gli stomi rimangono
quindi chiusi; in queste condizioni non c’è bisogno di
CO2 e quindi si conserva l’acqua. Anche durante il
giorno gli stomi si chiudono se la pianta sta perdendo
acqua a una velocità troppo elevata.
Figura 52. Gli stomi.
(A) Fotografia al microscopio elettronico a scansione di uno
stoma aperto delimitato da due cellule di guardia a forma di
salsiccia. (B) Meccanismo di apertura e chiusura degli stomi.
Le parole:
Stoma in greco significa «bocca», qui inteso nel senso
più ampio di «aperturta».

Le radici hanno funzione di ancoraggio al substrato e di assorbimento di acqua e minerali.
Nella maggior parte delle piante, l’acqua e i minerali
entrano attraverso ilsistema radicale, che è collocato nel
terreno (dove la luce non penetra); per questo motivo le
radici sono tipicamente prive di capacità fotosintetica.
Esistono principalmente due tipi di sistemi radicali.
Molte dicotiledoni possiedono un sistema radicale a
fittone: una singola grande radice primaria si accresce
in profondità nel terreno ed è accompagnata da radici
laterali meno pronunciate. Il fittone stesso svolge
spesso le funzioni di organo di riserva delle sostanze
nutritive, come accade nella carota (▶figura 53 A).
Le monocotiledoni e alcune dicotiledoni possiedono
invece un sistema radicale fascicolato, costituito da
filamenti che si propagano orizzontalmente
ramificandosi sotto la superficie del suolo (▶figura 53
B); poiché gran parte delle loro radici occupa i primi
centimetri del terreno, le monocotiledoni, in particolare
quelle erbacee, proteggono efficacemente il suolo
dall’erosione.
Figura 53. Due esempi di sistemi radicali.
Il sistema a fittone della carota (A) differisce dal sistema radicale
fibroso del porro (B).
Le piante utili per gli esseri umani.
Il ruolo svolto dalle piante nell’ambiente è fondamentale
e complesso, benefico anche per il mantenimento delle
risorse che ci sono utili. La vegetazione ha effetti
sostanziali sul suolo, sull’acqua, sull’atmosfera, sul
clima e sull’assetto idrogeologico.
I semi sono la nostra principale fonte nutritiva.
Tra le moltissime specie di piante con semi coltivate, ne
sono state individuate 12 che si ritengono basilari per la
nutrizione umana: riso, noce di cocco, frumento, mais,
patata, batata (o patata dolce), cassava (o tapioca o
manioca), canna da zucchero, barbabietola da zucchero,
soia, fagiolo e banana.
Oltre la metà della popolazione umana mondiale ottiene
gran parte delle calorie quotidiane dal prodotto di
un’unica pianta, il riso (Oryza sativa), particolarmente
importante nella dieta dei Paesi asiatici, dove è coltivato
da quasi 5000 anni. Le piante del riso sono usate in
molti modi, per esempio come rivestimento dei tetti,
come foraggio e lettiera per animali e anche per
confezionare abiti, e persino la lolla (il rivestimento del
seme scartato nella lavorazione) viene sfruttata in
diversi modi, da combustibile a fertilizzante.
In alcune popolazioni la noce di cocco (Cocos nucifera)
è chiamata «albero della vita», poiché, in effetti, tutte le
parti della pianta vengono in qualche modo utilizzate: il
tronco come materiale da costruzione, la linfa essiccata
come zucchero o fermentata come bevanda, le foglie
per rivestimenti di abitazioni, cestini e cappelli, i
germogli come verdura. Anche la noce di cocco, il
frutto, ha molti usi: il guscio duro si adopera come
contenitore o combustibile, lo strato fibroso mediano
per farne tappeti e imbottiture, mentre il seme,
costituito dal «latte» e dalla polpa, è usato come cibo e
bevanda (sterile).
Per milioni di persone la polpa del cocco è la principale
fonte di proteine; dalla polpa essiccata (commerciata
con il nome di copra) e pressata si ottiene l’olio di
cocco, l’olio vegetale più usato nel mondo nelle
industrie (lubrificanti, gomma sintetica) e, sebbene con
proprietà nutritive scarse, anche per l’alimentazione. La
copra si usa anche come fertilizzante e come mangime
per il bestiame.
Le piante sono usate come medicine
dall’antichità
Dalle piante si ricava una miriade di principi attivi che
permettono di curare dal semplice disturbo alla vera e
propria malattia. In passato, gli unici farmaci erano
quelli naturali e tra questi, i principali erano proprio
quelli estratti dalle piante. Una delle più antiche
«professioni» che si possono immaginare è quella del
cosiddetto «uomo della medicina» o sciamano (un ruolo
spesso ricoperto da donne), la persona che cura usando
rimedi derivati dalle piante. Oggi anche noi usiamo
molte medicine derivate da piante. Il campo della
ricerca di nuovi principi attivi di origine vegetale è
molto attivo e ha già permesso di scoprire farmaci
importanti nel trattamento di vari tipi di malattie.




![Gimnosperme 2012-2013 x pdf [modalità compatibilità]europa.uniroma3.it/biologia/documenti/b4a7b8d9-5a45-4cc9-ae4d... · Unica conifera spontanea in Europa caducifoglie Coni ♂globosi](https://static.documenti.site/doc/80x56/5badd8c309d3f2b5718b80a5/gimnosperme-2012-2013-x-pdf-modalita-compatibilita-unica-conifera-spontanea.jpg)


