

2
&Stato e valutazioneStato e valutazione
del metabolismodel metabolismo nell’obesitànell’obesità
Pietro Morini Sabrina Severi
RICERCACLINICA

Indice
Il bilancio energetico nell’uomo 3
Sabrina Severi
Dieta e metabolismo 38
Pietro Morini

Il bilancio energetico nell’uomo Sabrina Severi
Specialista in Scienza dell’Alimentazione, Dottoranda in Fisiologia dei Distretti - Università di Modena
Possiamo definire come bilancio energetico l’equilibrio esistente tra l’energia immagazzinata e l’energia spesa. L’energia in entrata, corrispondente all’energia derivata dagli alimenti, deve essere in equilibrio con l’energia in uscita, quella relativa alle quote per il mantenimento della dimensione corporea, per il ripristino delle componenti tessutali perse, per consentire l’attività fisica, motoria e ricreativa. Nel caso in cui l’introito calorico sia minore o maggiore della spesa energetica avremo rispettivamente perdita o aumento di peso e, quindi, variazioni nelle riserve energetiche corporee. L’introito energetico alimentare, mostrato in figura 1, è costituito da carboidrati, proteine e grassi. Come è noto la completa utilizzazione dei macro nutrienti ai fini energetici - ossidazione - richiede ossigeno, che viene trasportato alle cellule attraverso il sistema circolatorio. Al termine dei processi di ossidazione i prodotti finali del metabolismo vengono eliminati attraverso la sudorazione (evaporazione), la respirazione (acqua e CO2
) e la diuresi (acqua e urea). Vanno considerate anche le piccole perdite di energia nelle feci (cibi non digeriti, cellule derivate dalla desquamazione delle pareti delle mucose, batteri) e nella miscellanea dei tessuti demoliti (capelli, cute, flusso mestruale ecc.). L’introito dei nutrienti deve essere tale da bilanciare le perdite descritte affinché i depositi di energia e la massa corporea rimangano costanti.
FIGURA 1
Diagramma di flusso dell’energia nel corpo umano, da Heymsfield S.B. (1995).
ES rappresenta l’energia immagazzinata dal corpo sottoforma di riserve di grasso (F), di glicogeno (G), e di proteine (P).
P EI
F
CHO
O2
Efeci
Calore
CO2 H2O
Misc. Urea
G
F P
ES

4
Stato e valutazione del metabolismo nell’obesità
È altrettanto noto (tabella 1) che i principali depositi di energia del corpo sono rappresentati da grassi e proteine; l’altra piccola componente è rappresentata dal glicogeno, generalmente ignorata negli studi perché poco significativa dal punto di vista calorico e comunque difficilmente misurabile. È difficile stabilire il ruolo effettivo di ciascun nutriente nella formazione dei depositi corporei in quanto ognuno mostra interazioni con l’altro: gli aminoacidi possono essere convertiti a glucosio o acidi grassi; il glucosio può essere convertito a grasso (figura 1).
TEE-TDEE Il dispendio energetico totale (Total Energy Expenditure o Total Daily Expenditure Energy) viene definito dalla somma di tre diverse componenti: metabolismo basale, termogenesi e attività fisica. La sua valutazione nel-l’uomo è di notevole importanza in quanto ci dà la possibilità di stabilire le necessità “energetiche-nutrizionali”. Sulla base della TEE vengono infatti elaborati i fabbisogni nutrizionali della popolazione sana, intesi come livelli di energia derivata dal cibo per bilanciare il dispendio energetico individuale. Il concetto di bilancio energetico per l’uomo sano è codificato come quota calorica necessaria per mantenere costante la dimensione e la composizione corporea, per supportare l’attività fisica giornaliera e per garantire la salute nel lungo termine e, inoltre, per mantenere l’attività fisica necessaria nelle attività socialmente ed “economicamente” desiderabili. Quest’ultima definizione più moderna e olistica, in quanto espressione del fabbisogno energetico vincolato al benessere psicofisico individuale, viene proposta sia dall’OMS che dalla FAO e dalla Commissione Europea. È stato evidenziato il rapporto esistente tra TEE e il peso corporeo e il BMR; tuttavia l’uso di un fattore moltiplicativo del BMR per ottenere TEE, potrebbe essere fonte di un errore significativo nel calcolo del dispendio giornaliero sul singolo individuo. Numerosi studi, infatti, confermano l’assenza di una stretta relazione tra il TEE e la percentuale di massa grassa corporea.
TABELLA 1
Caratteristiche dei depositi corporei dei macronutrienti
Proteine Carboidrati Grassi
Pool corporeo (kg) 12 (0.06) * 0.7 12 - 18
Intake giornaliero (g) 100 300 100
% di riserva giornaliera 0.8 0.7 0.5 - 0.8
Soggetti non obesi, nella norma, con uno stile di vita alimentare di stampo occidentale * I valori fanno riferimento al pool corporeo degli amminoacidi liberi
5
Il bilancio energetico nell’uomo
Negli obesi, ad esempio, possono verificarsi le seguenti condizioni, tra loro contrastanti: ■ il dispendio energetico aumenta per l’incremento della massa magra
che deve supportare nel tempo l’incremento di peso. ■ il dispendio energetico diminuisce se correlato a individui con
elevate percentuali di FAT corporeo. ■ il dispendio energetico aumenta per consentire il “trasporto” di
individui con peso superiore alla norma. ■ il dispendio per l’attività fisica giornaliera diminuisce se l’obeso ha
uno stile di vita prettamente sedentario.
Una quota variabile della spesa energetica è data dall’attività fisica ed intellettuale. Più l’attività fisica e intellettuale è intensa, tanto maggiore sarà il consumo calorico (tabella 2). Le suindicate condizioni possono tra-
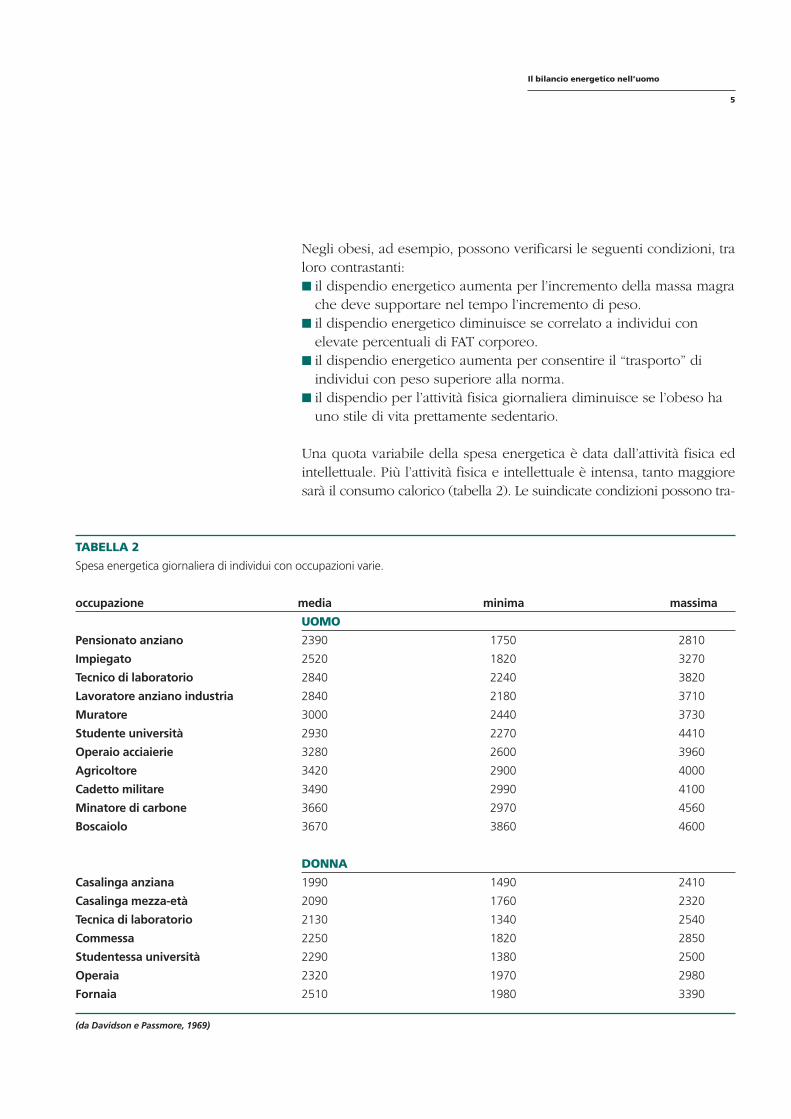
TABELLA 2
Spesa energetica giornaliera di individui con occupazioni varie.
occupazione media minima massima
UOMO
Pensionato anziano 2390 1750 2810
Impiegato 2520 1820 3270
Tecnico di laboratorio 2840 2240 3820
Lavoratore anziano industria 2840 2180 3710
Muratore 3000 2440 3730
Studente università 2930 2270 4410
Operaio acciaierie 3280 2600 3960
Agricoltore 3420 2900 4000
Cadetto militare 3490 2990 4100
Minatore di carbone 3660 2970 4560
Boscaiolo 3670 3860 4600
DONNA
Casalinga anziana 1990 1490 2410
Casalinga mezza-età 2090 1760 2320
Tecnica di laboratorio 2130 1340 2540
Commessa 2250 1820 2850
Studentessa università 2290 1380 2500
Operaia 2320 1970 2980
Fornaia 2510 1980 3390
(da Davidson e Passmore, 1969)

6
Stato e valutazione del metabolismo nell’obesità
dursi in valori sovrapponibili di TEE tra obesi e normopesi, o in valori differenti tra un obeso e l’altro, questi ultimi anche a parità di peso. Tutti questi aspetti rendono il calcolo teorico del TEE estremamente aleatorio, qualora lo si voglia applicare nella routine clinica sul singolo individuo; mentre lo rendono accettabile nella ricerca epidemiologica su gruppi selezionati per stato nutrizionale. Riassumendo: la spesa energetica totale (TEE) è data dalla spesa energetica basale (BEE), dall’energia spesa nell’attività fisica (AEE) e dalla termogenesi indotta dalla dieta (SDA):
TEE = BEE + SDA + AEE
Qui brevemente tracciamo la definizione delle tre componenti principali della TEE:
BMR-BEE Viene definito col termine di Metabolismo Basale (Basal Metabolic Rate o BMR) o spesa energetica basale (Basal Energy Expenditure o BEE), la quantità di energia spesa da un individuo nelle seguenti condizioni: ■ a digiuno da almeno 10-12 ore ■ in posizione supina e rilassata, tale da ridurre al minimo l’attività
muscolare ■ con temperatura corporea normale ■ a una temperatura ambiente neutrale (27-29°) per mantenere
l’omeostasi termica ■ in assenza di stimoli o stress psicologici e fisici.
Si devono quindi evitare rumori esterni che inducono stimolazioni al sistema nervoso autonomo o che provocano anche le più lievi contrazioni muscolari, evitare condizioni patologiche, o stati febbrili, e qualsiasi squilibrio termico. Il freddo infatti induce un aumento del tono muscolare e vasocostrizione, con conseguente maggior produzione di calore; mentre il caldo aumenta la vasodilatazione periferica, la sudorazione e l’attività cardiorespiratoria per consentire i meccanismi di termoregolazione.
I fattori che aumentano il BMR possono essere così riassunti: ■ età ■ sesso ■ temperatura corporea ■ peso corporeo ■ composizione corporea

7
Il bilancio energetico nell’uomo
■ fattori genetici ■ condizioni fisiopatologiche ■ ormoni
Massa magra, età e sesso sono i principali determinanti del metabolismo basale, poiché spiegano circa l’80% della variabilità inter individuale.
Età. La spesa energetica basale varia in funzione dell’età (LARN - tabella 3). Espressa come kcal m-2 hr-1, è massima alla nascita (53 kcal a 1 anno) e decresce fino ai valori minimi dopo i 70 anni (31 kcal a 75 anni). Se consideriamo invece la spesa calorica giornaliera (TEE - tabella 4), questa aumenta dalla nascita fino all’età dello sviluppo per raggiungere il massimo a 14-15 anni nelle femmine e 18-19 anni nei maschi e decresce col progredire degli anni. La diminuzione del metabolismo con l’età si correla con la diminuzione della massa magra e con un progressivo rallentamento del metabolismo energetico cellulare.
TABELLA 3
LARN. Equazioni di predizione del Metabolismo di Base (MB) a partire dal peso corporeo (Pc), espresso in kg, e, per bambini e adolescenti,
peso corporeo e statura (A) espressi in metri. Il metabolismo di Base risulta espresso in MJ/giorno.
età in anni MB (a partire dal peso) MB (a partire da peso e statura)
MASCHI
<3 0,2490 Pc - 0,13 0,0007 Pc + 6,35 A - 2,58
3-9 0,0950 Pc + 2,11 0,0820 Pc +0,55 A +1,74
10-17 0,0740 Pc + 2,75 0,0680 Pc + 0,57 A + 2,16
18-29 0,0640 Pc + 2,84
30-59 0,0485 Pc + 3,67
60-74 0,0499 Pc + 2,93
≥75 0,0350 Pc + 3,43
FEMMINE
<3 0,2440 Pc - 0,13 0,068 Pc + 4,28 A - 1,73
3-9 0,0850 Pc + 2,03 0,071 Pc + 0,68 A + 1,55
10-17 0,0560 Pc + 2,90 0,035 Pc + 1,95 A + 0,84
18-29 0,0615 Pc + 2,08
30-59 0,0364 Pc + 3,47
60-74 0,0386 Pc + 2,88
≥75 0,0410 Pc + 2,61
(dati derivati dal rapporto FAO/WHO/ONU -1985- e da Schofield et al. -1985-. Nell’anziano, i dati sono stati completati con studi italiani -Ferro-Luzzi, 1987-)
8
Stato e valutazione del metabolismo nell’obesità
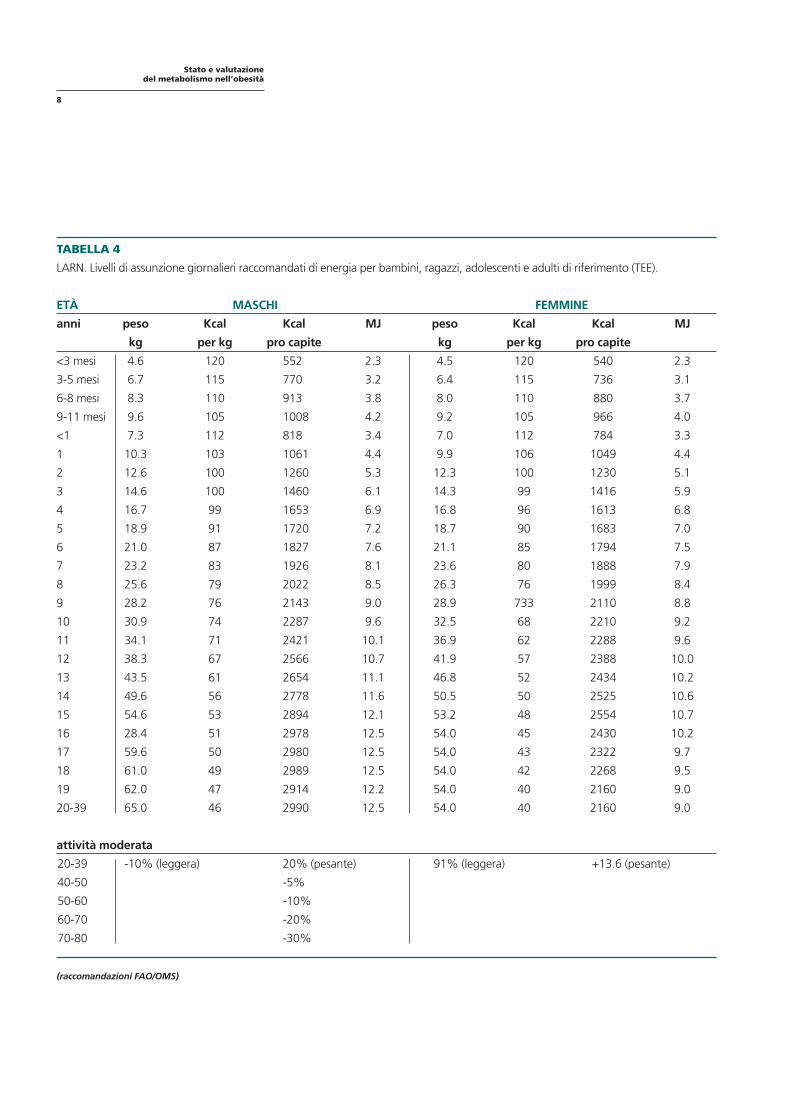
TABELLA 4
LARN. Livelli di assunzione giornalieri raccomandati di energia per bambini, ragazzi, adolescenti e adulti di riferimento (TEE).
ETÀ MASCHI FEMMINE
anni peso Kcal Kcal MJ peso Kcal Kcal MJ
kg per kg pro capite kg per kg pro capite
<3 mesi 4.6 120 552 2.3 4.5 120 540 2.3
3-5 mesi 6.7 115 770 3.2 6.4 115 736 3.1
6-8 mesi 8.3 110 913 3.8 8.0 110 880 3.7
9-11 mesi 9.6 105 1008 4.2 9.2 105 966 4.0
<1 7.3 112 818 3.4 7.0 112 784 3.3
1 10.3 103 1061 4.4 9.9 106 1049 4.4
2 12.6 100 1260 5.3 12.3 100 1230 5.1
3 14.6 100 1460 6.1 14.3 99 1416 5.9
4 16.7 99 1653 6.9 16.8 96 1613 6.8
5 18.9 91 1720 7.2 18.7 90 1683 7.0
6 21.0 87 1827 7.6 21.1 85 1794 7.5
7 23.2 83 1926 8.1 23.6 80 1888 7.9
8 25.6 79 2022 8.5 26.3 76 1999 8.4
9 28.2 76 2143 9.0 28.9 733 2110 8.8
10 30.9 74 2287 9.6 32.5 68 2210 9.2
11 34.1 71 2421 10.1 36.9 62 2288 9.6
12 38.3 67 2566 10.7 41.9 57 2388 10.0
13 43.5 61 2654 11.1 46.8 52 2434 10.2
14 49.6 56 2778 11.6 50.5 50 2525 10.6
15 54.6 53 2894 12.1 53.2 48 2554 10.7
16 28.4 51 2978 12.5 54.0 45 2430 10.2
17 59.6 50 2980 12.5 54.0 43 2322 9.7
18 61.0 49 2989 12.5 54.0 42 2268 9.5
19 62.0 47 2914 12.2 54.0 40 2160 9.0
20-39 65.0 46 2990 12.5 54.0 40 2160 9.0
attività moderata
20-39 -10% (leggera) 20% (pesante) 91% (leggera) +13.6 (pesante)
40-50 -5%
50-60 -10%
60-70 -20%
70-80 -30%
(raccomandazioni FAO/OMS)
9
Il bilancio energetico nell’uomo
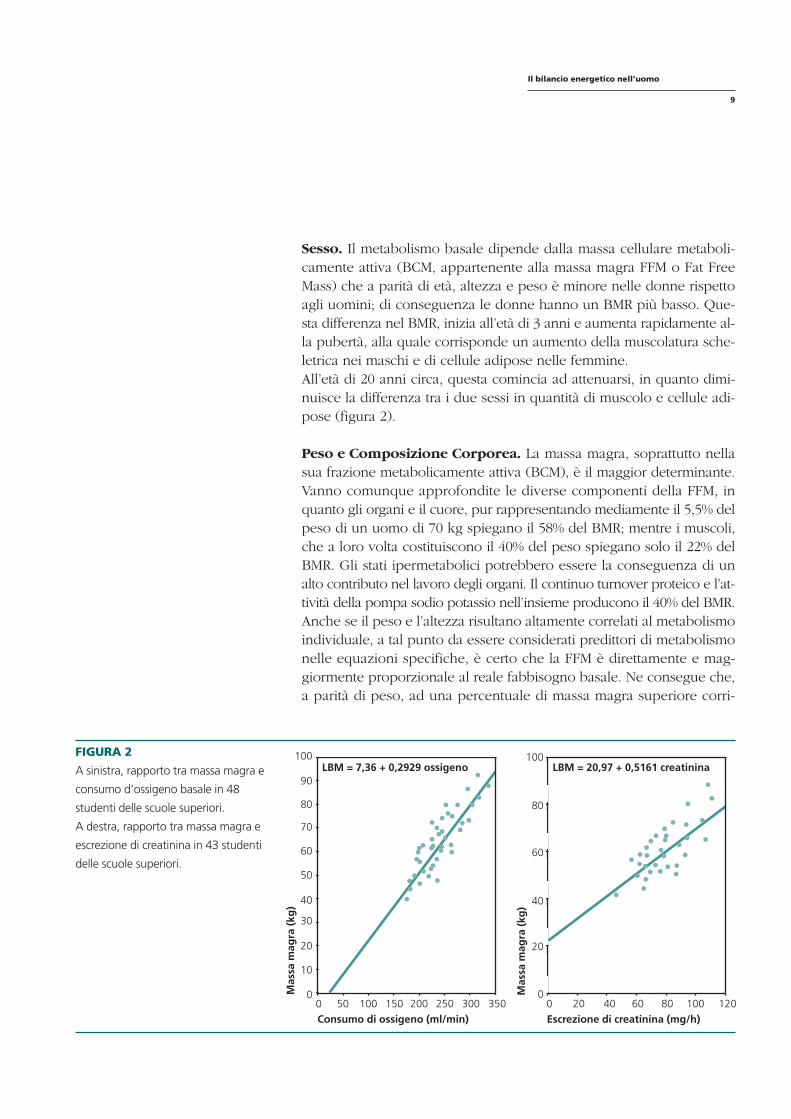
Sesso. Il metabolismo basale dipende dalla massa cellulare metabolicamente attiva (BCM, appartenente alla massa magra FFM o Fat Free Mass) che a parità di età, altezza e peso è minore nelle donne rispetto agli uomini; di conseguenza le donne hanno un BMR più basso. Questa differenza nel BMR, inizia all’età di 3 anni e aumenta rapidamente alla pubertà, alla quale corrisponde un aumento della muscolatura scheletrica nei maschi e di cellule adipose nelle femmine. All’età di 20 anni circa, questa comincia ad attenuarsi, in quanto diminuisce la differenza tra i due sessi in quantità di muscolo e cellule adipose (figura 2).
Peso e Composizione Corporea. La massa magra, soprattutto nella sua frazione metabolicamente attiva (BCM), è il maggior determinante. Vanno comunque approfondite le diverse componenti della FFM, in quanto gli organi e il cuore, pur rappresentando mediamente il 5,5% del peso di un uomo di 70 kg spiegano il 58% del BMR; mentre i muscoli, che a loro volta costituiscono il 40% del peso spiegano solo il 22% del BMR. Gli stati ipermetabolici potrebbero essere la conseguenza di un alto contributo nel lavoro degli organi. Il continuo turnover proteico e l’attività della pompa sodio potassio nell’insieme producono il 40% del BMR. Anche se il peso e l’altezza risultano altamente correlati al metabolismo individuale, a tal punto da essere considerati predittori di metabolismo nelle equazioni specifiche, è certo che la FFM è direttamente e maggiormente proporzionale al reale fabbisogno basale. Ne consegue che, a parità di peso, ad una percentuale di massa magra superiore corri-
FIGURA 2 100 A sinistra, rapporto tra massa magra e
90 consumo d’ossigeno basale in 48
80studenti delle scuole superiori.
0
20
40
60
80
100 LBM = 7,36 + 0,2929 ossigeno LBM = 20,97 + 0,5161 creatinina
Mas
sa m
agra
(kg
)
A destra, rapporto tra massa magra e 70
escrezione di creatinina in 43 studenti 60 delle scuole superiori.
50
40
Mas
sa m
agra
(kg
)
30
20
10
0 0 50 100 150 200 250 300 350 0 20 40 60 80 100 120
Consumo di ossigeno (ml/min) Escrezione di creatinina (mg/h)

10
Stato e valutazione del metabolismo nell’obesità
sponde un metabolismo basale più elevato. Contrariamente alle aspettative, nei soggetti obesi i valori di BMR risultano elevati se confrontati con quelli dei soggetti magri, dato che la quantità di FFM presente è a sua volta superiore. Ciò è in parte spiegabile dalle considerazioni sopra enunciate e cioè dall’aumento della massa magra che sempre accompagna l’eccessivo aumento del FAT. Poiché anche l’attività fisica incrementa la massa magra è buona norma non considerare il valore assoluto di BMR, ma il rapporto BMR/FFM (cal/kg) come sinonimo di efficienza metabolica individuale e come base per gli studi di ricerca e di clinica applicativa. Va inoltre ricordato che anche il tessuto adiposo ha un ruolo metabolico non trascurabile e questo alla luce delle recenti acquisizioni relative al tessuto adiposo bruno e bianco. In generale si ritiene che il tessuto adiposo nell’uomo adulto contribuisca per il 4-5% del BMR. Nei regimi fortemente ipocalorici il declino rapido del BMR appare strettamente correlato alla diminuzione della BCM, ma come vedremo più avanti trova spiegazioni anche in una ridotta termogenesi da dieta.
Temperatura Corporea. La temperatura corporea dell’uomo è costante, come in tutti gli animali omeotermi, ed è indipendente dalla temperatura ambiente. Un aumento della temperatura corporea dovuto a febbre, provoca un incremento della richiesta di ossigeno e aumenta il BMR. È stato calcolato un aumento di circa il 13% nei valori di metabolismo basale per ogni aumento di 1 grado di temperatura corporea; ma attenzione: questo incremento della temperatura corporea non nasce da influenze esterne, ovvero dagli scambi con l’ambiente, quanto dal lavoro metabolico interno di un organismo soggetto a ipertermia. La dimensione corporea incide nel fabbisogno basale anche attraverso la capacità di ritenzione termica individuale: un soggetto obeso, ad esempio, mostra per la sua maggiore superficie di tessuto adiposo sottocutaneo una maggiore ritenzione termica e quindi un minor dispendio energetico. Inoltre la tipologia del tessuto adiposo (adiposo bruno o adiposo bianco) costituisce l’elemento fondamentale nella conservazione o nella dispersione termica corporea. Anche la temperatura ambiente influenza in modo sensibile la spesa calorica. La temperatura alla quale si dice che il corpo umano è in “omeostasi termica” (27-29°C in assenza di vapore acqueo) è quella in cui si ha il minimo dispendio energetico. Quando la temperatura scende al di sotto del valore di neutralità termica (27°C), si verifica un aumento della spesa energetica come conseguenza della produzione del calore che va a compensare quello perso. Al di sotto dei 12°C il corpo non riesce più a compensare la perdita di calore; di conseguenza, rimanendo per un tempo sufficientemente lungo al di sotto di questa temperatura si ha

11
Il bilancio energetico nell’uomo
la morte per assideramento. Sopra i 29°C viene dissipata energia attraverso la sudorazione per mantenere la temperatura corporea a 37°C, di conseguenza si ha aumento della spesa energetica. Il corpo riesce a compensare temperature ambientali piuttosto elevate, fino a che la temperatura interna non comincia a salire. Quando quest’ultima raggiunge i 42°C circa si ha la morte dell’individuo. L’esercizio fisico prodotto in un ambiente sfavorevole (temperature superiori o inferiori ai 27°C) comporta quindi un maggior dispendio energetico, in quanto alla componente calorica dovuta al lavoro muscolare si aggiunge quella dovuta ai processi di termoregolazione corporea. Nell’ambito di un progetto di incremento del dispendio energetico per soggetti obesi, optare per un esercizio in un ambiente caldo piuttosto che per uno in ambiente freddo, non dovrebbe mutare l’ordine delle risultanze: in entrambi i casi il dispendio risulta aumentato. Tuttavia da alcune considerazioni legate al ruolo del tessuto adiposo bianco, come organo di conservazione o riduzione della dispersione termica, e ai fattori limitanti la termoregolazione in ambiente caldo, quali gli alti livelli di umidità, si dovrebbe arguire che l’esercizio in ambiente freddo porterebbe a maggiori dispendi energetici: esattamente il contrario delle proposte di mercato che promuovono cappe riscaldate, saune, infrarossi e simili.
Condizioni Fisiopatologiche. Tra le condizioni fisiologiche che inducono un lieve incremento del BMR per la necessità di supportare l’accrescimento “in toto” o di parti dell’organismo, sono presenti l’infanzia, l’adolescenza e la gravidanza, anche se in quest’ultima condizione numerosi lavori dimostrano un effetto compensatorio sul TEE per una ridotta termogenesi e attività fisica giornaliera. La fase mestruale rappresenta un periodo in cui il fabbisogno energetico basale può aumentare relativamente alle perdite ematiche e tessutali con fluttuazioni del 6-7 % sui valori basali; mentre la fase del sonno è caratteristica di un ridotto (il minimo giornaliero) fabbisogno energetico. Tra le condizioni patologiche che incrementano il BMR vi sono gli stati febbrili, lo scompenso cardiaco congestizio, l’ipertensione, le neoplasie, le malattie muscolo-scheletriche che inducono movimenti involontari. Tra le condizioni che riducono il BMR: l’anoressia nervosa, l’ascite, la malnutrizione calorico-proteica, i regimi fortemente ipocalorici (-15/25% del fabbisogno calorico giornaliero), alcune endocrinopatie come il morbo di Addison, la sindrome di Cushing.
Fattori genetici. Gli studi più suggestivi legati alla relazione tra la genetica e la spesa energetica sono stati ricavati dall’osservazione di una particolare etnia di indiani americani, i Pima, che secolarmente hanno

12
Stato e valutazione del metabolismo nell’obesità
vissuto in condizioni climatiche sfavorevoli con lunghi periodi di carestia. Tali condizioni hanno abituato l’organismo a tesaurizzare l’energia introdotta (Thrifty gene o gene della frugalità) a tal punto che l’adesione al costume occidentale più ricco in fonti alimentari ha prodotto una popolazione mediamente obesa e diabetica. Analizzando i Pima obesi, si è riscontrata la presenza di livelli di adiposità ascrivibili a precise aggregazioni familiari, suggerendo una caratterizzazione genetica con un metabolismo basale basso per quelli suscettibili a un maggior sviluppo di obesità. La disposizione dei distretti adiposi in altre etnie (boscimani e altre popolazioni africane e australi) suggerirebbero una stretta relazione tra metabolismo e riserve energetiche corporee, dimostrando una alta efficienza metabolica nell’immagazzinamento delle calorie ingerite. Studi recenti su modelli animali (ratti ob/ob) suggerirebbero l’esistenza di alcuni difetti genetici a livello del tessuto adiposo, causa di una riduzione nella produzione di leptina, o di una iperproduzione di leptina inefficiente, con conseguente iperfagia e riduzione del metabolismo basale. Trasportate tali esperienze sull’uomo, si è ipotizzato che l’obesità non sia tanto ascrivibile all’ipotesi leptino-priva quanto a una insensibilità recettoriale, o a una resistenza, nei confronti della leptina stessa. Un altro ceppo di topi (fat/fat) mostra avere un difetto metabolico diverso, legato alla mutazione del gene per la carbossipeptidasi, ma con le stesse risultanze di ridotto dispendio energetico basale. Inoltre, la constatazione che i genitori obesi tendono a generare figli obesi con una probabilità del 73% e che un solo genitore obeso mantiene una probabilità del 41% fa sorgere l’ipotesi che la componente genetica, e quindi metabolica, sia prevalente rispetto ai fattori alimentari, culturali e sociali. Esistono forme genetiche (sindromi di Prader-Willi, Laurence Moon-Biedl, Allstrom) causa di obesità infantile per le quali si possono ipotizzare anomalie nel metabolismo energetico. E infine: studi compiuti su coppie di gemelli monozigoti e dizigoti dimostrano che i fattori genetici sono responsabili del 40-50% della variabilità interindividuale nella termogenesi indotta dal cibo.
Cibo. L’introduzione del cibo determina a sua volta variazioni nella spesa energetica. Prove metaboliche hanno dimostrato che la spesa energetica aumenta dopo un pasto. Questo incremento del consumo energetico può essere considerato come il lavoro richiesto per il metabolismo dei nutrienti o il lavoro di digestione, e si esprime come DIT (Termogenesi Indotta dalla Dieta), o come SDA (Azione Dinamico Specifica). Il lavoro di digestione dipende dalla qualità e dalla quantità dei cibi ingeriti: grassi e carboidrati hanno una SDA bassa (5 e 5-10% rispettivamente), mentre quella delle proteine è relativamente alta (10-35%). Questi valori sono indicativi e dipendono dai differenti processi meta

13
Il bilancio energetico nell’uomo
bolici di volta in volta attivati: per esempio, la conversione del glucosio a glicogeno comporta una spesa corrispondente al 5% del contenuto energetico del glucosio, mentre sale a 24% se il glucosio è convertito in lipidi. Inoltre la DIT dipende dalla frequenza e dalla durata dei pasti, dalla dimensione e dalla palatabilità del cibo, dallo stato nutrizionale dei soggetti e da eventuali condizioni fisiopatologiche. Riguardo ai soggetti obesi, il consumatore di snack, o il “grignotteur” rappresenterebbero tipologie di comportamento alimentare di maggior rischio per le implicazioni metaboliche legate alla frequenza di consumo, alla palatabilità e qualità degli ingredienti – più zuccheri semplici e grassi – aventi un costo metabolico relativamente basso per la deposizione e un potere saziante altrettanto basso. Esattamente la soluzione ideale per l’industria alimentare che deve promuovere l’alto consumo.
Ormoni. Diversi ormoni influenzano il BMR, stimolandolo o rallentandolo, in quanto possono incrementare i processi di ossidazione dei substrati corporei ai fini energetici o all’opposto, possono promuovere i processi di sintesi. L’insulina ha un’azione ipoglicemizzante e promuove lipogenesi e glicogenosintesi, svolgendo quindi un’azione sia di stimolo che di risparmio energetico. Le variazioni ponderali dei soggetti con disfunzione tiroidea si spiegano con gli effetti degli ormoni tiroidei sul metabolismo: il BMR aumenta nell’ipertiroidismo e diminuisce negli stati di ipotiroidismo. Anche il calo ponderale rapido, derivato da iperfunzionalità tiroidea, risulta in parte dovuto a una riduzione della componente FFM e non alla componente FAT e quindi, l’efficienza metabolica è tale da ridurre la componente metabolicamente più attiva. Il paziente ex obeso si ritroverà con un BMR sostanzialmente ridotto. Ricordando l’influenza dell’ipertiroidismo sull’aumento dell’appetito, della temperatura corporea e della sensibilità ai recettori specifici alle catecolamine si evince come l’assetto ormonale vada a incidere su gran parte delle componenti metaboliche. Le catecolamine a loro volta aumentano la termogenesi, dimostrando che attraverso la stimolazione dei beta3 recettori si ottiene lipolisi con una maggiore dissipazione del calore; mentre l’NGF (fattore nervoso di crescita) sembra essere inversamente correlato all’attività del sistema simpatico. Infine, glucagone, testosterone e ormone somatotropo aumentano il BMR pur con meccanismi tra loro differenti. La sindrome dell’ovaio policistico, per esempio, si esprime clinicamente con iperandrogenismo rilevabile con concentrazioni elevate di testosterone e androstenedione, che coinvolgendo tutto l’assetto degli steroidi sessuali sarebbe causa di elevate anormalità metaboliche. La rilevazione su questi pazienti di obesità, ma soprattutto di grasso viscerale, potrebbe correlarsi a un effetto di liposintesi e quindi di risparmio energetico.

14
Stato e valutazione del metabolismo nell’obesità
RMR-REE Nelle condizioni definite basali, il BMR rappresenta l’energia che l’organismo utilizza per compiere il lavoro necessario al mantenimento del-l’omeostasi interna (sintesi e/o degradazione di alcuni costituenti cellulari, cicli biochimici, turnover proteico ecc.). In un individuo adulto con attività fisica media, il BMR rappresenta il 60-75% circa della spesa energetica giornaliera. Il BMR viene spesso confuso con il metabolismo a riposo (Resting Metabolic Rate o RMR o REE) che rappresenta l’energia spesa in condizioni di riposo. BMR e REE differiscono tra loro per la termogenesi indotta dalla dieta (diet-induced thermogenesis o DIT) conosciuta anche come Azione Dinamico Specifica dei nutrienti (Specific Dinamic Action o SDA); mentre non è ancora chiaramente espressa la differenziazione per gli altri effetti termogenici, quali la termogenesi indotta dalle variazioni termiche ambientali, da agenti come caffeina e nicotina e da stimoli psicologici, come ansia e paura. In sintesi l’equazione comunemente più accettata è:
REE=BMR + DIT
DIT-SDA-TEF L’incremento termogenico quindi, può essere suddiviso schematicamente in due componenti: termogenesi “obbligatoria” e “facoltativa”. La DIT, o SDA, o ancora TEF (Thermic Effect of Food), rientra nella componente obbligatoria con una piccola quota del consumo calorico giornaliero. In particolare, rappresenta l’energia spesa per la digestione, l’assorbimento e l’immagazzinamento dei nutrienti pari al 10-15% circa della spesa energetica totale. La termogenesi facoltativa, che partecipa per il 30-40% della DIT, comporta l’attivazione del sistema nervoso simpatico con meccanismi di difficile valutazione e oggi frutto di ricerca (figura 3). Il tipo di alimentazione può modulare il REE se l’alimento viene assunto poche ore prima della misura diretta o indiretta del metabolismo. L’influenza del pasto è tale che l’incremento del valore basale può presentare picchi differenti in funzione della qualità del cibo assunto, con un effetto che persiste anche a distanza di 5-6 ore dal pasto stesso. Un pasto a base di carne aumenta la DIT in misura maggiore (25-30%) rispetto ai carboidrati o ai grassi. È buona norma quindi, valutare la DIT complessivamente, e cioè durante e alla fine del periodo su indicato, in funzione della tipologia dell’alimento assunto. Studi ormai consolidati nel tempo (Apfelbaum 1969) dimostravano che il RMR diminuiva dell’1% al giorno nei soggetti a digiuno similmente a ciò che accadeva nei soggetti alimentati con 55 grammi di caseina, una quantità sufficiente a garantire un bilancio azotato in equilibrio. Studi più recenti (Prentice 1995) ipotizzano una diminuzione del 5% del RMR già al
15
Il bilancio energetico nell’uomo
primo giorno di dieta a 900 calorie. Appare evidente che il fabbisogno basale a riposo include una variabile legata all’alimentazione e che la sua riduzione nella dieta è legata a una ridotta SDA dato il pasto più povero e contenuto in quantità. Il declino del RMR nel digiuno può raggiungere un plateau del 25% in meno nell’arco di 3-4 settimane con un decremento calorico pari a 120 kcal al giorno. Il decremento calorico nel digiuno prolungato verrebbe completamente assegnato al costo del metabolismo proteico: in tali condizioni, per esempio, la sintesi proteica si ridurrebbe del 25% , mentre rimarrebbe inalterato il costo della ureogenesi e della produzione di ammonia. Rimandiamo ai paragrafi successivi l’approfondimento dell’influenza dei nutrienti sulla termogenesi.
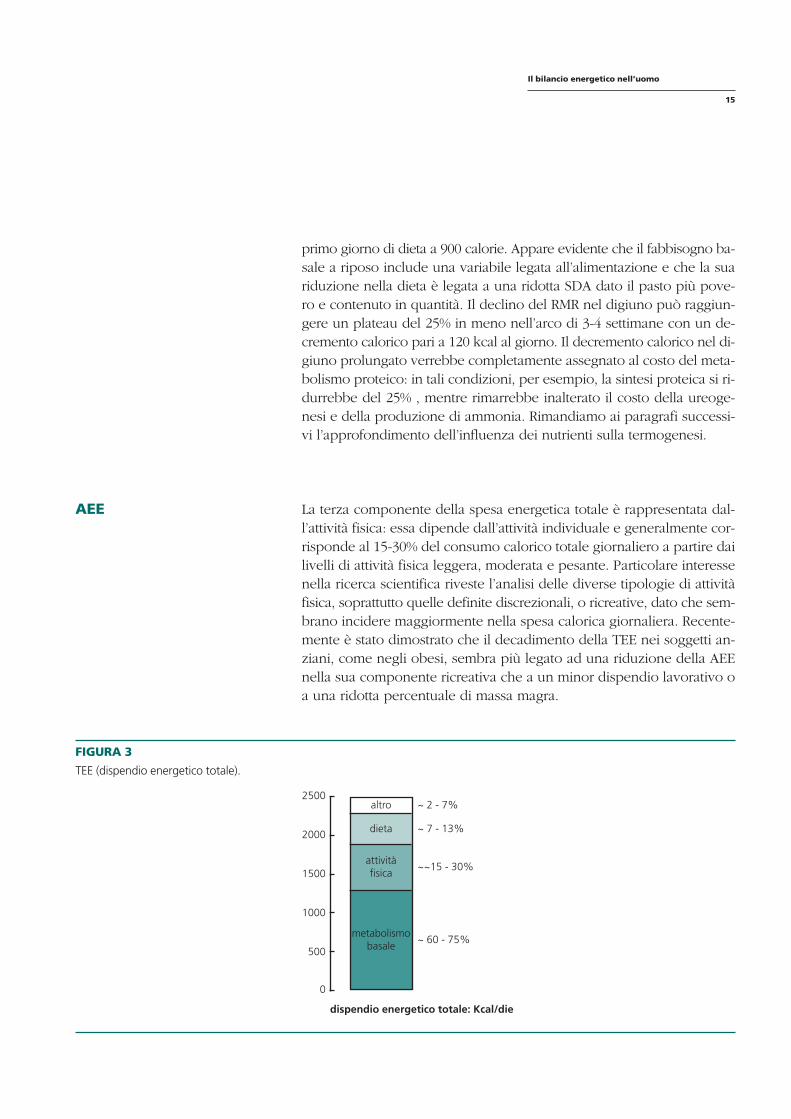
AEE La terza componente della spesa energetica totale è rappresentata dal-l’attività fisica: essa dipende dall’attività individuale e generalmente corrisponde al 15-30% del consumo calorico totale giornaliero a partire dai livelli di attività fisica leggera, moderata e pesante. Particolare interesse nella ricerca scientifica riveste l’analisi delle diverse tipologie di attività fisica, soprattutto quelle definite discrezionali, o ricreative, dato che sembrano incidere maggiormente nella spesa calorica giornaliera. Recentemente è stato dimostrato che il decadimento della TEE nei soggetti anziani, come negli obesi, sembra più legato ad una riduzione della AEE nella sua componente ricreativa che a un minor dispendio lavorativo o a una ridotta percentuale di massa magra.
FIGURA 3
TEE (dispendio energetico totale).
2500
2000
1500
1000
500
0
altro
dieta
attività fisica
metabolismo basale
~ 2 - 7%
~ 7 - 13%
~~15 - 30%
~ 60 - 75%
dispendio energetico totale: Kcal/die

16
Stato e valutazione del metabolismo nell’obesità
IL CALCOLO DEL METABOLISMO BASALE
Il consumo calorico viene misurato in kilocalorie (kcal o cal)) o anche in Joule (J). Viene definita kcal o grande caloria la quantità di calore necessaria per aumentare di 1°C (da 14.5 a 15.5 °C) la temperatura di 1 litro di acqua. Il principale mezzo di dispersione del calore dell’organismo è la pelle, appare perciò sensato rapportare la spesa energetica alla superficie corporea esprimendolo come kcal/m2. Du Bois e Du Bois condussero esperimenti su 8 uomini e 2 donne che permisero di misurare la superficie corporea. Da questi studi ricavarono una formula empirica per il calcolo della superficie corporea (SA)
SA= H0.725 x W0.450 x 71.84
dove H é l’altezza in cm, W è il peso in kg
Il lavoro di Du Bois e Du Bois per il calcolo della superficie corporea, pur essendo ancor oggi preso come riferimento, fu oggetto di critiche dato il basso numero di soggetti esaminati; Harris e Benedict, in seguito, proposero una formula per il calcolo della spesa energetica che impiegava variabili quali sesso, età, altezza e peso in base a studi condotti su 136 maschi, 103 femmine e 94 bambini. Numerosi studi sperimentali, condotti in comparazione con metodiche calorimetriche dirette e indirette, dimostrano l’attendibilità delle formule di Harris-Benedict con differenze sostanzialmente circoscritte intorno alle 50-100 kcal nei soggetti sani in normopeso. Tali formule sono:
BEE (kcal) uomini = 66.5 + 13.75W + 5.003H- 6.775A
BEE (kcal) donne = 655 + 9.563W +1.850H - 4.676A
Dove A = età in anni, W = peso in kg, H = altezza in cm.
Queste equazioni, pubblicate nel 1919, sono ancora utilizzate nella pratica clinica. Per i bambini, maschi e femmine, sono state formulate equazioni diverse: ■ per i bambini di età compresa tra i 6 e i 10 anni. BMR (kj/die) = 33.1 x peso (Kg) + 20.1 x altezza (cm) - 60.9 x età (anni) - 285 x sesso
(M = 0,F = 1) + 1542
■ per i bambini di età compresa tra i 10 e i 16 anni: BMR (Kj/die) = 50.2 x peso (Kg) + 29.6 x altezza (cm) - 144.5 x età (anni) - 550 x sesso
(M = 0,F = 1) + 594.3

17
Il bilancio energetico nell’uomo
È stato riscontrato che negli adulti i valori di BEE e REE calcolati con le formule, differiscono da quelli determinati attraverso l’analisi calorimetrica soltanto del 10% circa, quindi sono abbastanza attendibili. Queste formule possono essere usate solo su soggetti normopeso. In soggetti in cui sono presenti patologie quali l’obesità, i valori devono essere corretti attraverso funzioni popolazione-specifiche; inoltre, in soggetti in cui sono presenti infezioni gravi o ustioni, il metabolismo basale può incrementare anche del 100%, mentre in casi di malnutrizione, può diminuire fino al 40%. Il BMR misurato può essere confrontato con il valore considerato standard ed espresso come percentuale del rapporto con lo standard attraverso la formula:
(BMR misurato - BMR standard) x 100 BMR =
BMR standard
Il risultato viene espresso come la differenza percentuale dal valore predetto: è considerato nella norma il valore di BMR che si discosta dal predetto di ± 10%. Sempre all’inizio del secolo molti studiarono nuove formule per il calcolo teorico della spesa energetica, ottenendo risultati molto simili, come possiamo vedere: ■ 9 uomini normali (Du Bois)= valore medio 39.7 kcal/h-1m-2
m-2■ 9 uomini normali (Means)=valore medio 39.6 kcal/h-1
■ 82 uomini normali (Harris-Benidict)= valore medio 38.9 kcal/h-1m-2
Per la maggioranza dei soggetti normali, i valori di BEE (Basal Energy Expenditure) o REE (Resting Energy Expenditure) misurati effettivamente differiscono rispetto al valore predetta dal calcolo teorico di ± 10 %. La BEE può essere predetta in modo abbastanza preciso in gruppi di uomini e donne simili per età e composizione corporea partendo dal peso corporeo e dall’altezza. Il calcolo teorico della spesa energetica basale nei singoli individui varia rispetto alla stima effettiva misurata sul soggetto. La variabilità della BEE suggerisce che esistano delle differenze nell’efficienza metabolica tra i diversi individui. La tabella 5 da Flatt riporta la BEE in funzione della composizione corporea. Partendo da un ipotetico uomo adulto standard di 70 kg, vengono riportate diverse situazioni con scostamenti di ±40% rispetto al “peso ideale”. Viene riportata la BEE in funzione della superficie corporea e anche quella calcolata in funzione di variazioni della massa cellulare corporea o Body Cell Mass (BCM). Come si può notare, la differenza tra i

18
Stato e valutazione del metabolismo nell’obesità
valori della colonna A (calcolati in funzione della superficie corporea) e i valori della colonna B (calcolati in funzione della variazione della BCM), ammonta a ±15%. Ciò suggerisce che le variazioni della BEE nei diversi soggetti potrebbero essere attribuite a variazioni della BCM in rapporto al peso corporeo.
LA MISURA Esistono diversi metodi, o apparati strumentali, che vengono impiegati DELLE COMPONENTI nella pratica clinica per la misura delle componenti metaboliche. Per METABOLICHE brevità di descrizione possono essere raggruppati in metodi diretti e in
diretti. Si raccomanda per l’approfondimento della tecnica la consultazione della review di Prentice, Murgatroyd e Shetty, pubblicata sull’International Journal of Obesity, dal titolo: Techniques for the measurement of human energy expenditure – a practical guide.
La calorimetria diretta. La calorimetria diretta si basa sul principio per cui tutta l’energia consumata dall’organismo per compiere lavoro viene liberata sotto forma di calore. Pertanto, misurando la perdita di calore corporeo per evaporazione, con-
TABELLA 5
Dispendio energetico e variazioni nella composizione corporea.
53 (c) 60 (b) 70 (a) 84 (d) 98 (e)Peso (kg)
Percentuale peso ideale (%) 76 86 100 120 140
Scheletro (kg) 7 7 7 7 7
Compartimenti intercellulari (kg) 20 17.5 17.5 17.5 17.5
Tessuto adiposo (kg) 5 5 17.5 29.5 43.5
Massa cellulare (kg) 21 30 28 30 28
Superficie (m2) 1.66 1.75 1.87 (a) 2.02 2.16
BMR (Kcal/m2/h) 36.5 36.5 36.5 (a) 36.5 36.5
Dispendio energetico “A” (Kcal/die) 1450 1530 1640 1770 1890
Massa cellulare (Kcal/kg) 69 ! 51 ! 59 59 68 !
Dispendio energetico “B” (Kcal/die) (f) 1230 1760 1640 1760 1640
B-A/A (%) -15 +15 (0) -1 -13
(a) Ipotizzando un uomo adulto di 40 anni alto 178 cm – Geigy, Scientific Table, 7th edition, Ciba-Geigy, Basel, 1970, p. 537 e 540. (b) “Tipo attivo”: diminuzione del tessuto adiposo e un certo incremento della massa cellulare. (c) Individuo denutrito, con perdita di tessuto adiposo e/o massa cellulare, tendenza all’edema nutrizionale. (d) Individuo sovrappeso, con sensibile aumento del tessuto adiposo e aumento contenuto della massa cellulare. (e) Individuo obeso, con rilevante aumento del tessuto adiposo ma senza aumento della massa cellulare. (f) Ipotizzando che il dispendio energetico sia proporzionale al peso della massa cellulare e che corrisponda a 58,6 kcal/kg di massa cellulare come
nell’individuo normale.

19
Il bilancio energetico nell’uomo
duzione, irradiazione e convezione, si può risalire al consumo di energia. Solitamente la calorimetria diretta viene effettuata in una camera isolata, adiabatica, dove il soggetto viene esaminato nell’arco di 24 ore in condizioni, successivamente, di riposo, di lavoro e di attività ricreativa. Il calore rilevato è dato per i 3/4 circa da perdite per conduzione e radiazione. Per poter registrare la perdita di calore attraverso le componenti evaporative, occorre che l’aria in entrata nella camera metabolica sia condizionata a temperatura e umidità fisse. L’aria che esce dalla camera transita poi attraverso uno scambiatore di calore identico a quello in entrata, in modo da essere riportata nella camera alla temperatura e umidità iniziali. Il calore estratto dall’aria in uscita rappresenta il calore perso attraverso l’evaporazione, che va sommato alla quota di calore disperso in processi non evaporativi. Di uso più semplice sono i calorimetri a gradiente, in cui viene misurata la trasmissione del calore attraverso uno strato isolante, di forma, grandezza e proprietà termiche costanti. Le temperature, sulle due superfici dello strato isolante, dipendono dalla velocità di produzione e trasmissione del calore. La differenza tra le due temperature dello strato isolante, sulla superficie interna e su quella esterna, viene misurata in condizioni di equilibrio termico e viene correlata alla quantità di calore disperso dal corpo all’interno dell’isolante. La calorimetria diretta permette una stima precisa, accurata e diretta del calore prodotto; d’altra parte, la metodica presenta degli svantaggi: l’elevato costo e l’impossibilità di avere informazioni sui substrati corpo-rei effettivamente utilizzati.
La calorimetria indiretta. Più semplice e pratica da attuare è la calorimetria indiretta, che misura i gas respiratori: l’anidride carbonica pro-dotta (CO
2) e l’ossigeno (O
2) di un determinato volume di aria espirata.
La calorimetria indiretta si basa su due principi fondamentali: ■ la prima legge della termodinamica (Legge della conservazione
dell’energia), secondo la quale l’energia non può essere nè creata nè distrutta, ma solo trasformata. Questa legge può essere espressa con la seguente equazione:
∆E = dQ + dW + dR
∆E = variazione dell’energia chimica
dQ = calore liberato dal sistema
dW = lavoro meccanico fornito
dR = altre forme di energia liberate

20
Stato e valutazione del metabolismo nell’obesità
■ l’energia prodotta dall’ossidazione dei cibi nell’organismo è uguale a quella prodotta dalla loro combustione nella bomba calorimetrica. La differenza è rappresentata dal fatto che nell’organismo l’ossidazione avviene a 37°C grazie ad una serie di enzimi che catalizzano le reazioni, mentre nella bomba calorimetrica l’ossidazione è diretta ed avviene a pressioni e temperature elevate. In termodinamica non è tanto importante il tipo di reazione che avviene, ma solo gli stadi iniziali e finali (reagenti e prodotti). Il nutriente ossidato viene trasformato in anidride carbonica ed acqua come nella bomba calorimetrica. Per quanto riguarda grassi e carboidrati, i prodotti finali dell’ossidazione sono CO2
ed H2O sia
nell’organismo sia nella bomba calorimetrica:
Glucosio C H1 O6 + 6O → 6CO + 6H O + calore 6 2 2 2 2
Tripalmitina C H O6 + 72.5 O → 51 CO + 49 H O + calore 51 98 2 2 2
Differente è la situazione delle proteine. Nella bomba calorimetrica le proteine sono trasformate in CO , H O, SO e N mentre nel corpo umano
2 2 4 2
l’azoto proteico non è trasformato in N2 ma in urea.
Ovoalbumina: (ossidazione nel corpo umano) 2 C H N O S + 532 O → 433 CO + 67CON H + 6H SO + 269 H O + calore 250 409 67 81 3 2 2 2 4 2 4 2
(ossidazione nella bomba calorimetrica) 4 C H N O S + 1265 O →1000 CO + 12H SO + 806H O + 134N2 + calore 250 409 67 81 3 2 2 2 4 2
L’energia prodotta dalle proteine nell’organismo sarà minore rispetto quella prodotta nella bomba calorimetrica, con una differenza calcolata sperimentalmente di 1.35 kcal/g di proteine.
Il sistema a circuito aperto o total body. Due sono i sistemi di calorimetria indiretta: a circuito chiuso e a circuito aperto o total body. Nel primo sistema il soggetto respira ossigeno puro da un circuito chiuso e l’espirato viene reimmesso nel sistema dopo aver sottratto la CO
2. Nel
la calorimetria a circuito aperto, invece, il soggetto respira aria ambientale in una camera chiusa. La camera è fornita di letto, tavolo, sedia, TV e tutto quanto può essere necessario per rendere l’ambiente confortevole. Il cibo e le bevande sono forniti attraverso una finestra a tenuta, le feci e le urine vengono eliminate attraverso un’altra. L’aria che viene fatta entrare nella camera deve essere ben miscelata, e temperatura e pressione devono essere continuamente controllate rispetto a un gas standard a composizione conosciuta. Gli scambi respiratori del soggetto vengono misurati dall’effetto che hanno sulla composizione dell’aria

21
Il bilancio energetico nell’uomo
nella camera metabolica. La differenza in concentrazione di ossigeno e di anidride carbonica tra l’aria in entrata e quella in uscita rappresenta lo scambio respiratorio del soggetto, dal quale può essere calcolata la spesa energetica. L’accuratezza della calorimetria indiretta total body è limitata dal flusso ventilatorio nella camera e dalla composizione dei gas. Nella misura degli scambi respiratori si può ottenere un’accuratezza entro ±1%. La precisione inoltre è limitata dall’amplificazione dei ”rumori” generati dagli strumenti d’analisi dei gas, ma questi sono volume dipendenti e direttamente influenzati dalla grandezza della camera metabolica. In una camera con un volume di 10 m3 gli scambi respiratori nel-l’arco di 30 minuti vengono misurati con una precisione che si avvicina a 4 ml/min, che corrisponde a una variazione della spesa energetica pari all’1.5-2 %. La precisione aumenta con il tempo per il quale si protrae la misurazione. Sembra, infine, che la qualità dei risultati dell’analisi effettuata per diverse ore sia più strettamente influenzata dalla grandezza della camera che dalla ventilazione. La calorimetria indiretta total body si basa su principi semplici e può fornire con precisione e accuratezza, oltre al dato sulla spesa energetica, anche informazioni sui substrati utilizzati. Tra gli svantaggi si possono citare gli elevati costi di gestione, e il ricorso a un ambiente artificiale nel quale é difficile combinare altre metodiche invasive.
Il sistema a circuito chiuso. Per la calorimetria indiretta a circuito chiuso sono disponibili apparecchi portatili per la misura dei gas respiratori. Essi dispongono o di una maschera, attraverso la quale il soggetto respira, oppure di una canopy nella quale va inserita la testa. Negli apparecchi dotati di canopy una pompa crea un flusso costante. Il flusso viene espresso in l/min. L’O
2 consumato e la CO
2 prodotta sono poi misu
rati nell’aria ventilata come differenza tra l’aria inspirata e espirata. Esistono anche apparecchi, solitamente utilizzati negli ambulatori, sprovvisti del-l’analizzatore di CO
2. In questi strumenti, la raccolta dell’aria espirata per
la misura della spesa energetica avviene utilizzando la sacca di Douglas. Quest’ultima è un involucro impermeabile ai gas, solitamente con volume di 100 l, nella quale viene raccolta l’aria espirata dal soggetto durante il test. La sacca di Douglas viene utilizzata per misurare il metabolismo sia a riposo sia in attività, in questo caso assicurandola al corpo del soggetto. La sacca di Douglas pesa circa 6.5 kg e può dunque essere d’ostacolo durante l’attività fisica. Esistono però anche strumenti con peso circa di 3 kg, che quindi interferiscono meno sull’attività del soggetto. Ovviamente, non essendo misurata la CO2
, questi strumenti non danno informazioni sui substrati utilizzati. Si stima che l’accuratezza degli strumenti portatili e ambulatoriali oscilli tra il +4% e il -2%.

22
Stato e valutazione del metabolismo nell’obesità
L’APPLICAZIONE Dalle rilevazioni di O2 e CO
2 presenti nell’aria inspirata ed espirata, si ri-
DELLA METODICA sale all’ossigeno espirato e all’anidride carbonica prodotta. Definiti come IF il flusso inspiratorio ed EF il flusso espiratorio si possono calcolare VO
2 e VCO
2:
VO = (IF - EFO ) x VE2 2 2
VCO = (IFCO - EFCO ) x VE2 2 2
dove VE è il volume di aria espirata
Le concentrazioni di gas sono trasformate in condizioni STPD (Standard Temperature Pressure Dry) e corrette per la frazione nella miscela dei gas. Per risalire al consumo energetico basale, esistono diverse equazioni (tabella 6) che mettono in rapporto il consumo di ossigeno e la produzione di anidride carbonica con il consumo calorico. Le equazioni sono basate sulla misura del consumo di O
2 (VO
2), della produzione di ani
dride carbonica (VCO2) e dell’escrezione urinaria, quest’ultima come sti
ma dell’entità dell’escrezione urinaria. Esse presentano la seguente forma (tabella 5):
kcal/die = a VO2 + b VCO2 – c N
in cui a, b e c sono coefficienti che dipendono dal tipo di substrato ossidato. Infatti, poter misurare in “vivo” la quota di grassi, proteine e carboidrati ossidati è il vantaggio principale della calorimetria indiretta. L’illustrazione di questo punto richiede di introdurre alcuni concetti chiave.
Il Quoziente Respiratorio. Il rapporto tra anidride prodotta ed ossigeno consumato (VCO
2/VO
2) è definito quoziente respiratorio (respi
ratory quotient o QR). Il QR differisce per i diversi substrati energetici: ■ lipidi, QR 0.7
TABELLA 6
Equazioni proposte per il calcolo del dispendio energetico: (Kcal/die = aVO2 + bVCO2 - cN).
Referenza a b c Kcal/die
Wair (1949) 3.94 1.11 2.10 2417
Consolazio (1963) 3.78 1.16 2.98 2347
Brower (1965) 3.87 1.20 1.43 2428
Frayn (1983) 3.76 1.18 0.88 2371
Brockway (1987) 3.96 1.08 1.41 2422
Jequier 3.84 1.09 0.61 2376

23
Il bilancio energetico nell’uomo
■ carboidrati, QR 1 ■ proteine, QR 0.83 ■ alcol, QR 0.66
Siccome ci sono quattro substrati e soltanto due variabili misurabili (ossigeno e anidride carbonica), per il calcolo della spesa energetica occorre conoscere le proteine e l’alcool ossidati (se quest’ultimo è stato consumato), che devono essere stimati indipendentemente. Per le proteine solitamente viene misurato l’azoto urinario, che rappresenta il prodotto finale dell’ossidazione proteica. L’ossidazione dei grassi e dei carboidrati può essere stimata attraverso gli scambi respiratori con una accuratezza rispettivamente pari a ±9.5 g/die (corrispondenti circa a 90 kcal) e a ±20 g/die (corrispondenti a circa 80 kcal). Una alimentazione corretta e bilanciata si caratterizza per un quoziente respiratorio compreso nel range 0.80 - 0.86.
Valori di QR al di fuori di questo range e possono essere spiegati come segue:
QR>1
Lipogenesi: è la trasformazione degli zuccheri in lipidi. Per esempio, trasformazione del glucosio in acido stearico:
3(C6H12O6) → C18H36O2 + 8O2
Nella lipogenesi si liberano 8 molecole di ossigeno, di conseguenza durante tale processo si ha un aumento di produzione di CO
2, mentre i va
lori di O2 consumato si mantengono relativamente stabili. Questo spie
ga perché il QR con valore attorno a 1 sia caratteristico della lipogenesi. È inoltre possibile che i valori di QR siano superiori se la VCO
2 aumenta
in modo rilevante. Generalmente lipogenesi e lipolisi avvengono simultaneamente. In alcuni tessuti la lipogenesi può essere caratterizzata da un QR inferiore a 1 se l’ossidazione dei grassi è quantitativamente più rilevante. Nel lavoro muscolare intenso e breve, si incrementa la produzione di acido lattico, spostando l’anidride carbonica dei bicarbonati della riserva alcalina all’espirato con un conseguente aumento del QR. Un quadro di iperventilazione del paziente, dovuto per esempio al fatto di dover respirare con una mascherina o un boccaglio, fa sì che il QR aumenti fino a valori superiori ad 1.
QR< 0.7

24
Stato e valutazione del metabolismo nell’obesità
LA STANDARDIZZAZIONE NELLA MISURA DEL METABOLISMO BASALE (BMR)
Esistono due condizioni in cui il QR ha valori inferiori a 0.7: gluconeogenesi e chetogenesi. La gluconeogenesi è la conversione di lipidi e proteine, molecole povere di ossigeno, a glucosio che invece è ricco di ossigeno. Anche in presenza di gluconeogenesi, spesso si hanno condizioni in cui il glucosio formato è successivamente ossidato per cui non se ne ha accumulo. Perchè il glucosio possa essere accumulato è necessario che la dieta sia ricca di proteine e povera di carboidrati. In queste condizioni, dopo un pasto si ha stoccaggio di glucosio sotto forma di glicogeno. Anche la chetogenesi (tipica del digiuno prolungato o del diabete) comporta la trasformazione di sostanze povere di ossigeno in sostanze ricche di ossigeno. L’ulteriore ossidazione di queste sostanze non è contemplata nell’equazione della calorimetria indiretta. La quantità di corpi chetonici nelle urine del soggetto può però essere usata per correggere i valori di O2
e CO2.
È necessario innanzitutto che venga predisposto un ambiente in cui possano restare solo il paziente e il medico operante. Questa stanza deve essere in penombra e deve essere priva di oggetti che possano distrarre o attirare l’attenzione del paziente, in modo da limitarne l’attività mentale. Il soggetto verrà fatto sdraiare su di un lettino in posizione supina, in modo che si rilassi il più possibile. Quando si ritiene che il paziente sia perfettamente rilassato e a suo agio, si applicano il boccaglio respiratorio, o la mascherina o ancor meglio la canopy, in modo che i gas espirati siano raccolti e convogliati ai rispettivi analizzatori. Solo una piccolissima quota di gas espirati viene convogliata agli analizzatori, mentre il resto, viene rimandato all’esterno del calorimetro nell’aria ambientale. Anche per questo motivo è necessario che al momento dell’esame ci siano poche persone nella stessa stanza, in quanto con la respirazione viene aumentata la CO
2 presente nell’ambiente e, dato che il calorimetro prende come ri
ferimento le concentrazioni dei gas nell’ambiente come costante, verrebbe alterato l’esito dell’analisi. Prima di raggiungere gli analizzatori, i gas raccolti transitano in un flussimetro che misura il volume respiratorio. Il gas espirato viene di norma analizzato ogni minuto (esistono apparecchi che possono adottare intervalli inferiori, per esempio 20 secondi), per calcolare il dispendio calorico riferito alle 24 ore. Generalmente durante una analisi metabolica non tutti i valori misurati possono essere usati per determinare il BMR. Accade spesso che, nei primi minuti del test, il paziente non sia perfettamente rilassato, oppure che nel circuito interno via sia ancora dell’aria ambiente. Gli strumenti più sofisticati sono in grado di segnalare i dati più attendibili (quelli in Steady State o stato di equilibrio),

25
Il bilancio energetico nell’uomo
LA DETERMINAZIONE DELLA SPESA ENERGETICA (EE)
cioè i valori che non si discostano l’uno dall’altro più di 50 - 100 kcal. Per esempio, vengono eliminati dal referto finale i valori di VO
2, VCO
2 e QR
che mostrano oscillazioni maggiori al 10%. Se lo strumento non è in grado di fare questa selezione, l’operatore deve controllare che i QR siano abbastanza costanti, o permettere ad esempio solo le oscillazioni inferiori al 5%, e controllare che i valori di metabolismo mostrino una certa costanza per almeno 5 minuti. Se non si riesce a raggiungere questa condizione in 30 minuti si rende necessario ripetere il test.
La spesa energetica può essere determinata in due modi: attraverso VO2
e VCO2 o solo con VO
2.
Determinazione con VO2 e VCO
2. Nell’adulto i volumi respiratori clas
sici sono: 0.250 l/min per ossigeno, 0.225 l/min per anidride carbonica e 0.01 g/min per azoto con una spesa energetica di 1.233 Kcal/min. Un errore del 10% nella misurazione dell’ossigeno comporta un errore del 7% nella spesa energetica, mentre un errore del 10% nella misurazione dell’anidride carbonica produce un errore del 3% e un errore del 100% nella valutazione dell’azoto crea un errore solo del 1%. In base a questi risultati sono state costruite equazioni per calcolare la spesa energetica:
EE (kcal/die) = 3.581 VO2 (l/die) + 1.448 VCO2 (l/die) - 32.4
EE (kcal/die) = 3.581 VO2 (l/min) + 1.448 VCO2 (l/min) - 0.022
Queste formule sono state usate in soggetti ammalati per valutare la differenza rispetto a quelle che prendevano in esame anche l’azoto. L’errore calcolato era di 27 kcal/die. Anche in questo caso è stato chiaramente dimostrato che la determinazione dell’azoto escreto non è necessario per il calcolo della spesa energetica, anche nei pazienti in cui la secrezione di azoto è largamente variabile.
Determinazione con VO2. La misura più importante per determinare
la spesa energetica è quella della VO2. In alcune circostanze si può omet
tere la CO2 cosi come l’azoto. Come già si è detto, un errore di VCO
2
pari al 10% provoca un errore del 3% nella EE. Questo significa che se il QR può essere stimato con un’approssimazione del 10% così VCO
2
può essere calcolato da VO2 con la medesima approssimazione.
Generalmente in una popolazione normale il QR non ha scostamenti dal valore di 0.9 superiori a ±10%, fatta eccezione per condizioni tran

26
Stato e valutazione del metabolismo nell’obesità
METABOLISMO CIRCADIANO E BIOCHIMICA DEI NUTRIENTI
sitorie, per le quali è possibile trascurare la misura di VCO2. Per malati
il cui QR varia da 0.7 a 1.2, si rende necessaria la misura di entrambi i gas. Al calcolo della EE con il solo ossigeno si ricorre di norma durante l’esercizio o in fisiologia respiratoria:
1.0548 - 0.0504 EFO2EE (kcal) = x VE 1 + 0.28 p
5.04EE (kcal) = x (0.2093 - EFO ) x VE
1 + 0.082 p 2
EFO2 = concentrazione dell’ossigeno nell’aria espirata
VE = volume espiratorio p = frazione dell’energia totale della dieta derivante dalle proteine Il valore (0.2093 - EFO
2) rappresenta la differenza nel contenuto di os
sigeno tra l’aria inspirata e l’espirata.
VCO2 = VO2 x QR
EE (kcal) = 3.581 VO2 + 1.448 QR x VO2 - 1.773 Nu
Come abbiamo indicato nei paragrafi precedenti, il dispendio energetico del metabolismo basale viene ricavato sulla base del consumo di ossigeno a digiuno (almeno 8 ore dall’ultimo pasto) e quindi si preferisce effettuare il test al mattino. Anche la qualità del pasto può influire sul quoziente respiratorio e, quindi, si preferisce ridurre la componente proteica, così che il quoziente respiratorio sia determinato quasi esclusivamente dalle componenti glucidiche e lipidiche. Il minimo dispendio energetico nelle 24 ore si osserva durante il sonno notturno e corrisponde al 10 % della quota relativa alle condizioni basali. Quando il soggetto è inattivo durante il giorno ma il suo metabolismo è stimolato a intermittenza dal consumo di cibo e dalla termogenesi, la sua spesa energetica é solitamente più alta del 20% rispetto alle condizioni basali (figura 4). Quest’ultimo valore, ripartito nelle 24 ore, descriverebbe più compiutamente il REE, che sommato a sua volta al dispendio per l’attività fisica porterebbe al dispendio energetico totale nella giornata. Permane tuttavia, il quesito riguardante la misurazione delle diverse componenti metaboliche nella realtà quotidiana, dato che è possibile solo simulare il dispendio del soggetto in esame nella camera metabolica, simulando lo sti
27
Il bilancio energetico nell’uomo
le di vita; mentre l’effettiva attività potrebbe essere significativamente diversa per l’incidenza delle relazioni sociali e dei fattori ricreativi e ludici. Occorre sottolineare che questi fattori comportano di per sé un incremento del BMR legato alla componente catecolaminergica. Come abbiamo riferito nei paragrafi precedenti gli aumenti nella spesa energetica durante il giorno dovuti all’assunzione di cibo vengono assegnati alla SDA (Specific Dynamic Action of Food). La SDA varia a seconda dei nutrienti: 2-7% per i carboidrati, 20-30% per le proteine, 2-4% per i lipidi. Per una dieta mista il valore di SDA usualmente utilizzato é pari il 10%. Questo aumento del dispendio energetico è dovuto essenzialmente al costo energetico per il trasporto e la conversione dei nutrienti assorbiti nelle rispettive forme d’immagazzinamento. La qualità del pasto, quindi, varia sensibilmente la SDA. Poiché l’ossidazione dei substrati comporta produzione di ATP, la misura del dispendio energetico fornisce anche la misura dell’ATP prodotto. Per queste ragioni “dispendio energetico” e “produzione di ATP” sono termini spesso interscambiabili (tabella 7). Il valore di 7 kcal per l’idrolisi di una mole di ATP é quello generalmente utilizzato. Da un punto di vista fisiologico però non ha significato, perché tale valore varia a seconda della concentrazione dei diversi reagenti che partecipano alle reazioni. Nei globuli rossi, per esempio, la concentrazione di ATP e ADP é tale che l’energia prodotta per l’idrolisi di una mole di ATP corrisponde a 12.9 kcal/mole (Lehninger 1975). Dall’altra parte la spesa energetica per la
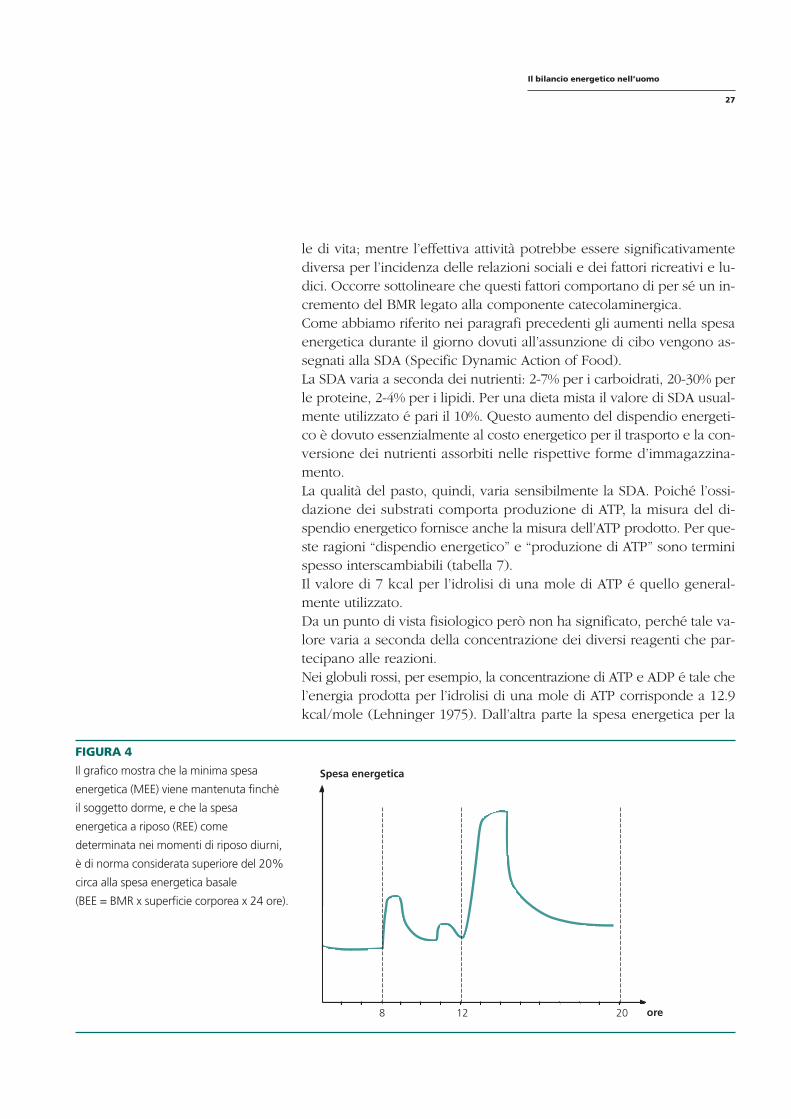
FIGURA 4
Il grafico mostra che la minima spesa Spesa energetica
energetica (MEE) viene mantenuta finchè
il soggetto dorme, e che la spesa
energetica a riposo (REE) come
determinata nei momenti di riposo diurni,
è di norma considerata superiore del 20%
circa alla spesa energetica basale
(BEE = BMR x superficie corporea x 24 ore).
8 12 20 ore

28
Stato e valutazione del metabolismo nell’obesità
sintesi di una mole di ATP dovrebbe essere basata sul calore di combustione di una mole di substrato diviso il numero di ATP generati da questa ossidazione. Questo valore si aggira a 18-20 kcal/mole. Questo significa che una spesa energetica giornaliera di 2400 kcal implica che 2400/18 = 133 moli di ATP vengano idrolizzate e rigenerate giornalmente. Nel paragrafo successivo verranno ripresi i concetti di SDA per i processi digestivi e di Energia Dissipata per i processi di deposizione dei nutrienti come riserve corporee.
IL DESTINO METABOLICO Nella figura 5 vengono riportate le diverse vie metaboliche previste per DEI NUTRIENTI l’amido ingerito con la dieta. I numeri indicano le moli di ATP formato
(+) e utilizzato (-) per ogni mole di glucosio. La differenza tra ATP formato e ATP utilizzato esprime la quantità di energia immagazzinata sotto forma di ATP: ATP GAINED. L’utilizzazione e la produzione di ATP nello schema metabolico, indica un bilancio tra il numero di “ATP gained” (ATP guadagnato) e “ATP made” (ATP formato) entrambi derivanti dagli amidi della dieta, o dal glicogeno endogeno, o dalle proteine o dai trigliceridi presenti come depositi corporei. Il trasporto attivo del glucosio nell’intestino è presumibilmente legato all’estrusione del sodio, con un consumo di 1 mole di ATP ogni 3 atomi di Sodio pompato fuori dalle cellule epiteliali (Crane 1962). Il costo in ATP per il coinvolgimento degli enzimi intestinali nel trasporto e nella mobilizzazione del glucosio dall’intestino al torrente ematico viene arrotondato a 0.5 ATP; mentre per la produzione di glicogeno è di 2 ATP e per il ciclo di Cori di 1 ATP.
TABELLA 7
Ossidazione del substrato glucidico.
Substrato
1 Glucosio
1 Palmitato
1 AA
4.5 Glucosio
Ossigeno necessario
6 O2
23 O2
5.1 O2
4 O2
Prodotti dell’ossidazione
6 CO2 + 6 H2O
16 CO2 + 16 H2O
4.1 CO2 + 0.7 urea + 2.8 H2O
1 Palmitile + 11 CO2 + 11 H2O
Energia prodotta (Kcal)
673 (a)
2398 (a)
475 (b)
630 (c)
QR
1.0
0.70
0.80
2.75
(a) Handbook of Chemistry and Physics, 51st edition, 1970, p. D 217 (b) I coefficienti stechiometrici per l’ossidazione delle proteine variano leggermente in funzione della composizione in aminoacidi. L’esempio prodotto
qui si riferisce all’ossidazione di 100 grammi di bistecca di bovino adulto (McGilvery, 1970). Il calore della combustione è stato calcolato usando un valore di 4,32 kcal/g di proteina (Geigy Scientific Tables, 1970, p. 539).
(c) (4,5 moli Gluc x 673 kcal/mole) – 2398 kcal/mole Palmitato = 630 kcal. La stechiometria della reazione, comprese la prosuzione e l’utilizzazione di legami ad alta energia (~), è basata su quella riportata per la conversione del glucosio in grasso nel tessuto adiposo del ratto (Flatt, 1970).

29
Il bilancio energetico nell’uomo
Poiché l’ATP per l’assorbimento, il trasporto e il deposito del glucosio si assume abbia un equivalente calorico di circa 18-20 kcal per ogni mole utilizzata, la SDA% degli amidi ingeriti può essere ricavata dividendo le calorie da ATP impiegate nel processo digestivo per le calorie forni-
FIGURA 5
La via metabolica.
ossidazione -2 ATP -1ATP -2 ATP -0,5 ATP
ciclo di cori -1 ATP -1ATP -1 ATP FFA riciclo -1,6 ATP
PALMITICO fegato
- 0.5 ATP
AMIDO (162 g x 4.15 cal/g)
GLUCOSIO (1 mole = 673 Kcal)
intestino
GLICOGENO fegato muscoli
PALMITICO tessuto adiposo
GLUCOSIO sangue
CO CO CO CO2 2 2 2
BIL
AN
CIO ATP utilizzato
ATP formato
ATP guadagnato
-5,5
+38
32,5
-4,5
38
33,5
-3,5
38
34,5
-10,8
38
27,2
ATP utilizzato
per l’assorbimento,
il trasporto
e la deposizione
SDA
2,5
7%
2,5
7%
0,5
2%
8,7
26%
energia dissipata
per la deposizione 5% 3% 19%

30
Stato e valutazione del metabolismo nell’obesità
te da una mole dell’amido stesso. Ad esempio, la SDA% nel caso del glicogeno epatico, sarà di 2,5 ATPx20 kcal divise per 673 kcal e cioè più del 7%. Nello schema: se il glucosio viene tutto immagazzinato nel sangue la SDA% è pari al 2% circa; se viene convertito in glicogeno la SDA% è pari al 7%, se però tutto l’amido viene trasformato e depositato come grassi la SDA% sale al 26% circa. Ciò corrisponde a una condizione sfavorevole per la lipogenesi da carboidrati; ma se per esempio, il nostro pasto si presentasse lievemente eccedente il fabbisogno per una quota energetica del 20%, l’eccedenza si trasformerebbe in grassi con una SDA% complessiva, risultante dalla quota di glicogeno e di trigliceridi depositati, pari al 10%. La riduzione della SDA in questo caso è l’espressione di una condizione favorevole per la lipogenesi non tanto per il ruolo metabolico dei carboidrati di per sé, quanto per l’ipercaloricità del pasto in quel momento. In ultimo, viene riportata l’energia dissipata per la deposizione del glucosio, sotto forma di riserva corporea, che oscilla dal 3-5% se si tratta di glicogeno sino al 19% se trigliceridi. L’energia dissipata è la differenza tra la SDA osservata quando il glucosio assorbito è direttamente ossidato, o quando esso è convertito in glicogeno o grassi prima dell’ossidazione. Costi irrisori nel contesto ossidativo o di glicogenesi, a dimostrazione del ruolo dei carboidrati come fonte di energia pronta e disponibile; costi elevati invece qualora si tratti di lipogenesi. In cifre, il costo metabolico, ossia l’energia dissipata per la conversione del glucosio a glicogeno è pari al 5% del contenuto energetico del glucosio, mentre sale al 19 % quando viene convertito a grasso. È d’obbligo una riflessione: l’organismo nel regolare la deposizione, o al contrario la mobilizzazione e ossidazione dei substrati, si esprime attraverso la sua capacità di gestire le calorie introdotte da un pasto all’altro al fine di sopperire alla richiesta energetica. Questo è vero, ma come vedremo più avanti, non significa che l’eventuale eccedenza calorica di un pasto sia immagazzinata come riserva estensibile di glicogeno o di proteine. Piuttosto, l’unico comparto corporeo estensibile è il tessuto adiposo e la risposta immediata all’eccedenza energetica del pasto è la deposizione dei grassi. Una volta depositati, il costo di mobilizzazione e ossidazione dei grassi, sempre tra un pasto e l’altro, è così elevato che preferenzialmente verranno intaccati i glucidi di riserva. L’ipercaloricità del pasto è quindi il fattore più pericoloso per la potenziale lipogenicità ma è anche, nella pratica dietetica, la più difficile da combattere. Infatti, la dieta ideale dovrebbe prevedere un’equa ridistribuzione dei pasti in ambito circadiano e la riduzione in termini calorici del pasto previsto alla sera, ma questo si confronta con la modificazione dello stile di vita del paziente, intervento ben più arduo di quello meramente educativo. Nella figura 6 vengono schematizzate le diverse vie metaboliche che possono seguire i grassi ingeriti con la dieta. I nu
31
Il bilancio energetico nell’uomo
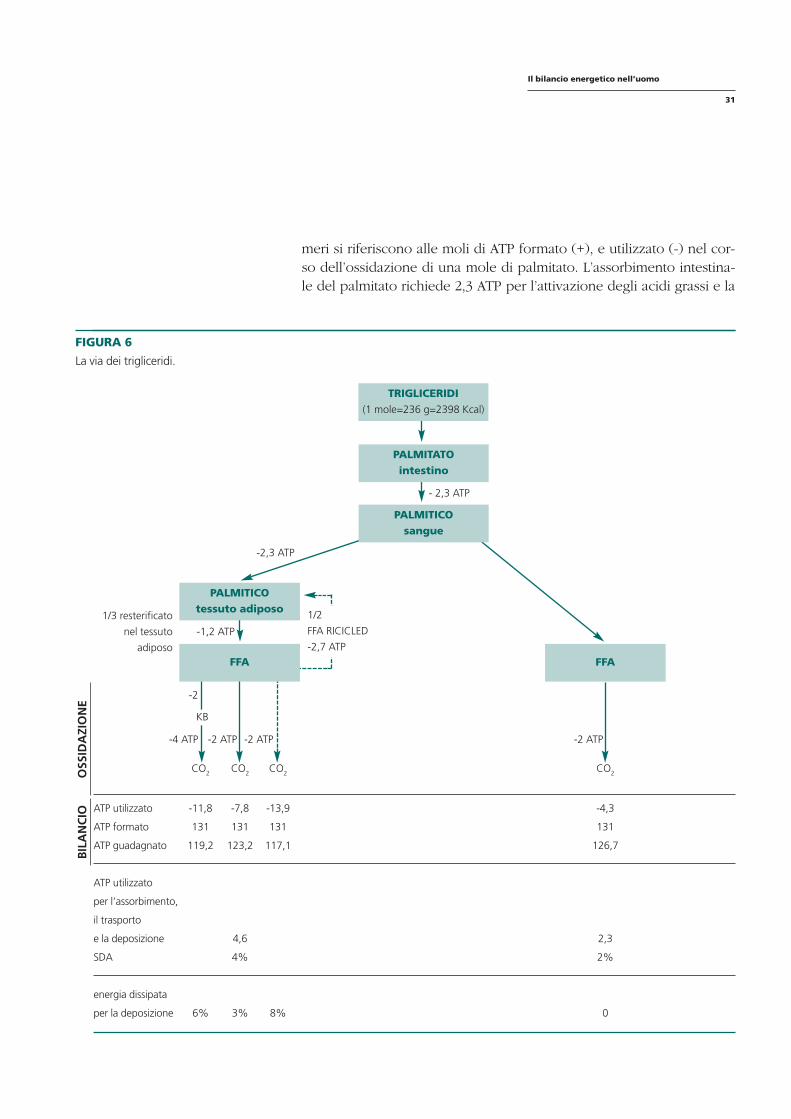
FIGURA 6
La via dei trigliceridi.
meri si riferiscono alle moli di ATP formato (+), e utilizzato (-) nel corso dell’ossidazione di una mole di palmitato. L’assorbimento intestinale del palmitato richiede 2,3 ATP per l’attivazione degli acidi grassi e la
- 2,3 ATP
TRIGLICERIDI (1 mole=236 g=2398 Kcal)
PALMITATO intestino
-2,3 ATP
1/3 resterificato
nel tessuto
adiposo FFA
-2 ATP
-2
-4 ATP -2 ATP -2 ATP
-1,2 ATP
PALMITICO tessuto adiposo
PALMITICO sangue
FFA
1/2
FFA RICICLED
-2,7 ATP
KB
BIL
AN
CIO
O
SSID
AZI
ON
E
CO2 CO2 CO2 CO2
ATP utilizzato
ATP formato
ATP guadagnato
-11,8
131
119,2
-7,8
131
123,2
-13,9
131
117,1
-4,3
131
126,7
ATP utilizzato
per l’assorbimento,
il trasporto
e la deposizione
SDA
4,6
4%
2,3
2%
energia dissipata
per la deposizione 6% 3% 8% 0

32
Stato e valutazione del metabolismo nell’obesità
loro esterificazione. La stessa quota di ATP è richiesta per il deposito nel tessuto adiposo (Ball, 1965). Quando gli acidi grassi liberi sono convertiti in corpi chetonici, un legame altamente energetico è utilizzato (GTP) per mole di corpi chetonici attivati in periferia. Le linee tratteggiate mostrano il riciclo e la riesterificazione degli acidi grassi liberi che sono in relazione con la spesa in ATP: viene assunto che un terzo degli acidi grassi prodotti nel tessuto adiposo è riesterificato in questo stesso tessuto, inoltre la metà degli acidi grassi nel plasma viene captata dal fegato, esterificata e rimessa in circolo come trigliceridi legati alle lipoproteine, con un costo di 2,7 ATP per acido grasso. La somma alla fine dello schema riporta la produzione e l’utilizzazione di ATP che era usata per calcolare l’ATP guadagnato e l’ATP formato dai grassi della dieta o endogeni. La SDA nel caso dei grassi alimentari si stima intorno al 2-4% in relazione agli acidi grassi direttamente ossidati o depositati. L’energia dissipata per la deposizione, a sua volta, oscilla dal 3 all’8%. Anche questi costi irrisori giungono a dimostrazione dell’estrema facilità dei grassi a depositarsi. La dieta dovrà essere di conseguenza essenzialmente ipolipidica! La figura 7 riporta le vie metaboliche degli aminoacidi ingeriti con la dieta. I numeri riportati mostrano le moli di ATP utilizzati(-) e formati (+) per 110 g (1 mole) di aminoacidi contenuti in una bistecca. (Mc Gilvery,1975). Il costo per il trasporto attivo degli aminoacidi nell’intestino non può essere stimato con precisione (Matthews, 1975), ma viene considerata mezza mole di ATP per aminoacido (ricordiamo, che come per l’assorbimento dei monosaccaridi, l’assorbimento degli aminoacidi è legato al-l’estrusione del sodio). Il costo per la sintesi di proteine è un costo elevato - 5,5 ATP- costituito da 4 ATP per la formazione del legame peptidico, 1 ATP per la sintesi di RNA e 0,5 ATP per il processo di assunzione degli aminoacidi stessi. Gli aminoacidi liberati dalla scissione delle proteine poi, vengono in parte riutilizzati per la sintesi di altri aminoacidi come si può vedere nella linea tratteggiata. L’ATP prodotto e utilizzato, corrisponde alle proteine della dieta e/o a quelle endogene. È da notare che la SDA è la stessa quando gli aminoacidi sono convertiti in proteine o direttamente ossidati, perché il costo della gluconeogenesi e ureogenesi è simile a quello della sintesi di proteine. Non tutti gli aminoacidi sono convertiti in proteine: sembra più verosimile che solo un terzo di quelli assunti con il pasto venga convertito, mentre due terzi vengano ossidati nelle ore successive il pasto stesso e, per sopperire al fabbisogno nei processi di risintesi, si riutilizzino gli aminoacidi derivati dal breakdown proteico.
33
Dieta e metabolismo
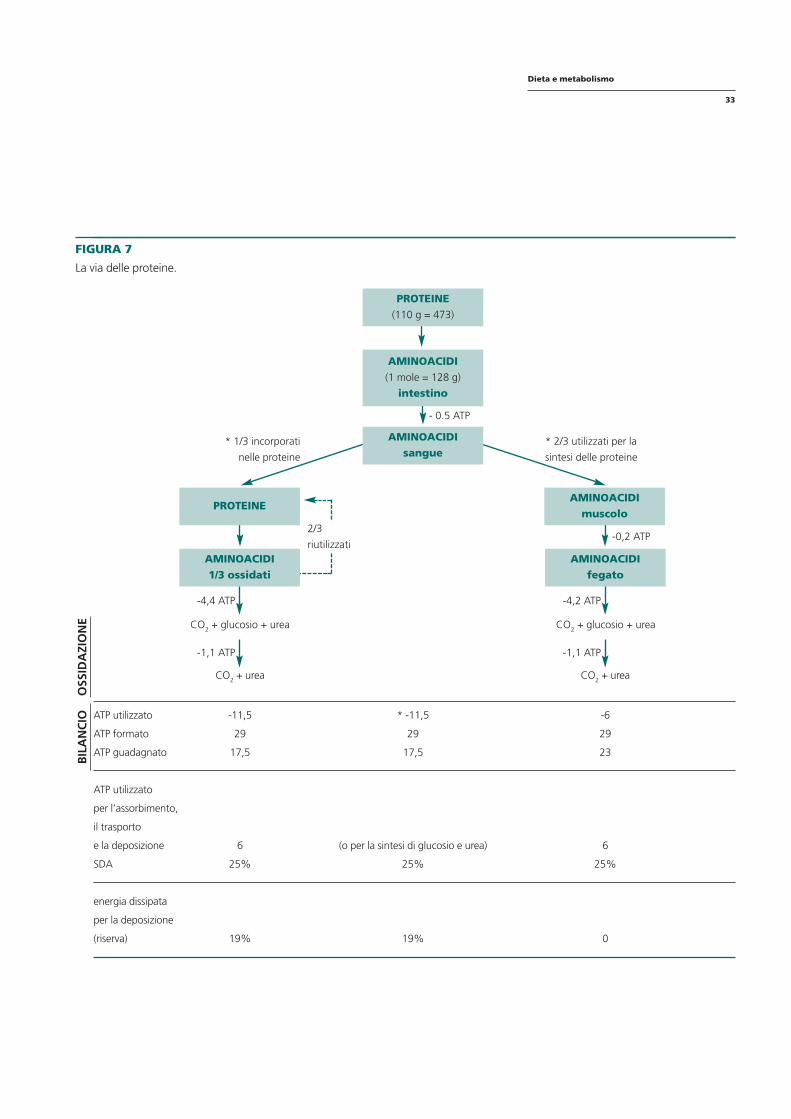
FIGURA 7
La via delle proteine.
AMINOACIDI fegato
* 1/3 incorporati
nelle proteine
* 2/3 utilizzati per la
sintesi delle proteine
-4,2 ATP
-0,2 ATP
PROTEINE AMINOACIDI
muscolo
AMINOACIDI 1/3 ossidati
2/3
riutilizzati
-4,4 ATP
- 0.5 ATP
PROTEINE (110 g = 473)
AMINOACIDI (1 mole = 128 g)
intestino
AMINOACIDI sangue
BIL
AN
CIO
O
SSID
AZI
ON
E CO2 + glucosio + urea CO2 + glucosio + urea
CO2 + urea
-1,1 ATP
CO2 + urea
-1,1 ATP
ATP utilizzato
ATP formato
ATP guadagnato
-11,5
29
17,5
* -11,5
29
17,5
-6
29
23
ATP utilizzato
per l’assorbimento,
il trasporto
e la deposizione
SDA
6
25%
(o per la sintesi di glucosio e urea)
25%
6
25%
energia dissipata
per la deposizione
(riserva) 19% 19% 0

34
Stato e valutazione del metabolismo nell’obesità
Le proteine producono la maggior quantità di ATP in rapporto alle calorie presenti nelle stesse proteine ingerite. Tuttavia, quando gli aminoacidi vengono convertiti in proteine, solo il 20% dell’ATP generato dall’ossidazione degli aminoacidi stessi viene utilizzato; un altro 20% dell’ATP generato deve coprire la gluconeogenesi e l’ureogenesi e quindi, solo il 60% dell’ATP formato nel corso dell’ossidazione riguarda il guadagno netto per l’organismo. Con una SDA così elevata (25%) e un ATP gained così ridotto si capisce perché la dieta iperproteica sia molto più ipocalorica di quanto lo dicano le calorie estrapolate dalle tabelle di composizione.
CONCLUSIONI Analizzando il rapporto tra ATP gained e ATP formato nei tre schemi metabolici si osserva che: ■ è 0,91 quando il glucosio derivato dall’amido ingerito viene
direttamente ossidato. ■ è 0,86-0,88 quando il glucosio viene convertito in glicogeno o per il
fegato o per i muscoli. ■ è 0,72 quando viene convertito in acidi grassi e poi ossidato, con
una energia dissipata pari al 19%. Inoltre: ■ l’energia dissipata per la deposizione del glicogeno dai carboidrati
della dieta è pari al 3-5 %. ■ l’energia dissipata per la deposizione dei grassi introdotti con la
dieta è pari al 8% a conferma dell’alta capacità di deposizione dei grassi
TABELLA 8
Calcolo dei costi dei nutrienti di stoccaggio.
Nutrienti Forma
di stoccaggio
Costo della sintesi Costo causato
dal riciclaggio
Energia dissipata
dallo stoccaggio
Carboidrati Glicogeno (fegato) 5% - 5%
Glicogeno (muscolo) 5% - 3%
Tessuto adiposo 24% 4% (a) 28%
5% (a)Grassi Tessuto adiposo 2% 7%
Proteine Proteine 24% - 24%
8% (b) 16% (b) 24%
(a) Ipotizzando gli FFA formati nel tessuto adiposo siano ri-esterificati per un terzo e per il 50% siano rilasciati dal tessuto adiposo per essere riciclati. (b) Ipotizzando che gli AA ingeriti siano convertiti in proteine solo per un terzo, ma che i due terzi degli AA liberati nel breakdown delle proteine siano
riutilizzati nella sintesi proteica.

35
SIGLE
Il bilancio energetico nell’uomo
■ l’energia dissipata per la deposizione delle proteine della dieta è pari al 19% a conferma della relativa difficoltà di formare riserve proteiche.
E per concludere: ■ quantitativamente il processo metabolicamente più costoso è la
conversione dei carboidrati ingeriti in riserva corporea di grassi (tabella 8). La SDA% relativa alla trasformazione dei carboidrati in grassi corporei e alla loro successiva ossidazione è più del 26% delle calorie inizialmente ingerite.
■ il processo metabolico meno costoso è l’assorbimento e la deposizione dei grassi della dieta a riprova che il modello dietetico più corretto è essenzialmente la dieta ipolipidica.
■ il processo più costoso è la deposizione delle proteine della dieta o la loro ossidazione a riprova dell’inutilità dei modelli iperproteici.
La strategia dietetica nella terapia dell’obesità appare allora codificata in un modello nutrizionale strettamente ipolipidico, lievemente ipoglucidico e normoproteico. Si dovrà invece, porre maggiore attenzione anche al-
ADP Adenosin Difosfato
AEE Activity Energy Expenditure
ATP Adenosin Trifosfato
BCM Massa Cellulare o Massa Metabolicamente Attiva
BCM Body Cell Mass
BEE Basal Energy Expenditure
BMR Basal Metabolic Rate
DIT Diet-inducet thermogenesis
EE Energy Expenditure
EF Flusso espiratorio
EFO2 Concentrazione di ossigeno nell’aria espirata
FFM Fat Free Mass
IF Flusso Inspiratorio
J Joule
Kcal Kilocaloria
QR Quoziente Respiratorio
RMR Resting Metabolic Rate
SDA Specific Dinamic Action
VCO2 Volume di anidride carbonica
VE Volume Espiratorio
VO2 Volume di ossigeno

36
Stato e valutazione del metabolismo nell’obesità
la piccola eccedenza calorica in quanto origine di pericolosi shift metabolici verso la liposintesi. Mentre i modelli iperproteici dissociati, alternati, aglucidici, alipidici o gli altri che ancora oggi vanno di moda, contrastano la biochimica dei nutrienti e il destino metabolico dei medesimi, non garantendo nel lungo termine l’ideale gestione delle riserve corporee.
BIBLIOGRAFIA 1. PR Murgatroyd, PS Shetty and AM Prentice. “Tecniques for the measurement
of human energy expenditure: a pratical guide”. International Journal of Obesity
(1993); 17:549-568
2. Marjolein Visser, Paul Deurenberg, Wija A van Staveren and Joseph Gaj
Hautvast. “Resting metabolic rate and diet - inducet Thermogenesis in young
and elderly subjects: relationship with body composition, fat distribution, and
physical activity level”. Am. J.Clin.Nutr. 1995; 61: 772-8
3. Daphne LE Pannemans and Klaasr Westerterp. “Energy expenditure, phisical
activity and basal metabolic rate of elderly subjects”. British Journal of Nutrition
(1995); 73:571-581
4. Susan B Roberts, Paus Fuss, Melvin Bheyman and Vernon R Young.
“Influence of age on energy requirements”. Am. J.Clin.Nutr. 1995; 62 (suppl.):
1053S - 8S
5. James O. Hill, Christopher Melby, Susal L Johnson and John C Peters.
“Physical activity and energy requirements”. Am J.Clin.Nutr. 1995; 62 (suppl.):
1059S -66S
6. Steven B Heymsfield, Pamella C Dariby, Lauren S Muhlheim, Dympna
Gallagher, Carla Wolper and David B Allison. “The calorye: myth,
measurement and reality”. Am. J.Clin.Nutr. 1995; 62 (suppl.): 1034S - 41S
7. Maffeis C, Schutz Y, Micciolo R, Zoccante L and Pinelli L. J. “Restin metabolic
rate in six - to ten - year - old obese and nonbese childern”. Pediatr. 1993; 122:
556-62
8. Molnar D, jeges S, Erhardt E and Shuts Y. J. “Measures and predicted resting
metabolic rate in obese and nonbese adolescents”. Pediatr. 1995; 127: 571-7
9. Simon Bursztein MD, David H Elwin PhD, Jeffrey Askanazi MD, John M
Kinney MD. “Energy Metabolism, Indirect Calorimetry and Nutrition”. Ed.
William & Wilkins - Baltimore Maryland - USA
10. AT Miller, JR and Carl S. Blyth. “Estimation of Lean Body Mass and Body Fat
From Basal Oxigen Consumption and Creatinine Excretion”.
11. Consolazio CF, Johnson RE, Pecora LJ. “Physiological measurement of
metabolic functions in man”. (1963) - McGraw-Hill, NY
12. Weir JB. “New methods of calculating metabolic rate, with special reference to
protein metabolism”. J Physiol 1949; 109, 1-9

37
Il bilancio energetico nell’uomo
13. Brower E “Report of sub-committee on constant and factors, in 3rd Symposium
on Energy metabolism”. European association for animal production Publ. N. 11,
Blaxer KL (ed). Accademic Press. London, 1965; pp 441-443
14. Frayn KN “Calculation of substrate oxidation rates in-vivo from gaseous
excange”. J.Appl.Physiol 1983; 55, 628-634
15. Brockway JM “Derivation of formulae used to calculate energy expenditure in
man”.. Hum.Nutr.Clin.Nutr. 1987; 41C, 463-471
16. Jequier E, Felber JP “Indirect calorimetry”. Clin. Endocr. Met. 1987; 1, (4), 911
935

Dieta e metabolismo Pietro Morini
Direzione Scientifica Centro Ricerche Dietosystem, Milano
Ritornando alla prima legge della termodinamica, abbiamo che l’energia introdotta nell’organismo sotto forma di alimenti (EI) deve essere uguale all’energia persa (EL) più l’energia immagazzinata come riserva (ES) (figura 1). Questa visione dell’alimentazione come “flusso di energia” enfatizza il concetto di dieta come fonte calorica necessaria a mantenere la massa cellulare, le funzioni dell’organismo, la salute e la longevità. L’energia immagazzinata è rappresentata dalla massa corporea e quando entrambe sono costanti vuol dire che siamo in perfetto bilancio energetico: ciò che viene introdotto bilancia esattamente ciò che viene perso. Si stima che ogni variazione di peso pari a un grammo abbia un equivalente calorico di 7,7 calorie. In realtà un bilanciamento così preciso rappresenta un fenomeno assai poco realistico, in quanto il mantenimento del corpo, in peso e composizioni stabili e costanti, comporterebbe un perfetto bilancio aritmetico tra l’energia introdotta e l’energia spesa nelle sue varie forme. Un’eccedenza di EI del 5% sul fabbisogno di un uomo di riferimento pari a circa 100 Kcal, ovvero un bicchierino di brandy, o 25 grammi di formaggio, o 30 g di pane, tanto per citare alcuni equivalenti alimentari, portano ad un incremento di 6-7 Kg di peso in un periodo di 5 anni. È dimostrato che la metà dell’incremento ponderale av-
FIGURA 1
La figura illustra il flusso energetico nel corpo.
Il perfetto bilancio energetico si ottiene quando l’energia immagazzinata e la massa corporea sono costanti.
EI ES EL EI = Energy Intake (introito energetico) ES = Energy Storage (riserve di energia) EL = Energy Loss (energia persa)

39
Dieta e metabolismo
MACRONUTRIENTI E RISERVE CORPOREE
viene già nel primo anno e che al termine del periodo considerato, il soggetto tende a riguadagnare peso per il raggiungimento di un nuovo “steady state”. La spiegazione potrebbe risiedere nella composizione del peso accumulato: ben il 75% è costituito da grasso, cioè dalla massa metabolicamente meno attiva. Ciò risulta confermato nella routine clinica quotidiana, ove si osserva che per una data eccedenza calorica l’obeso guadagna più peso rispetto al normopeso magro. Purtroppo le piccole variazioni nell’EI sono difficilmente misurabili, soprattutto negli obesi che tendono a sottostimare il proprio consumo alimentare per un’istintiva propensione alla gratificazione del medico intervistatore, come è stato recentemente sottolineato da Steven Heymsfield in un sua ricerca condotta all’Obesity Center Unit della Columbia University di New York. È quindi indispensabile nella pratica clinica stabilire il miglior mezzo di indagine alimentare in funzione della tipologia del paziente in esame e non adeguarsi a una metodologia standard di rilevazione. Questo impone un inquadramento preliminare del paziente per individuarne il profilo nutrizionale, in particolare per tipizzare il paziente nel suo rapporto con il cibo in, “restrained, unrestrained o disinibito”, e per valutarne le pulsioni e le motivazioni all’intervista; tutto questo prima ancora di stabilire il metodo di indagine alimentare. Si cerca di evitare in tal modo, il rischio di reports errati per i fattori psicosociali sopracitati. Purtroppo la prassi è quella di estendere a tutti i soggetti, indipendentemente dal loro profilo nutrizionale il metodo analitico ritenuto più preciso o, comunque, più facile nell’uso a seconda della propria esperienza personale. Il profilo nutrizionale e l’analisi del consumo alimentare, supportata da immagini di cibi e piatti, di volumi e porzioni, sono i due step procedurali che, se adottati in sequenza, dovrebbero ridurre l’errore nella stima dell’EI anche a livello della clinica ambulatoriale. Al di là delle perplessità legate alle metodologie per la rilevazione dell’effettivo EI, gli studiosi oggi tendono a misurare le componenti del dispendio energetico giornaliero (TEE), forse perché ritenute deducibili con maggior precisione e, inoltre, di maggior interesse clinico.
L’introito energetico è costituito da carboidrati, proteine e grassi. Come è noto la completa utilizzazione dei macronutrienti ai fini energetici - ossidazione - richiede ossigeno, che viene trasportato alle cellule attraverso il sistema circolatorio. Al termine dei processi di ossidazione i prodotti finali del metabolismo vengono eliminati attraverso la sudorazione (evaporazione), la respirazione (acqua e CO
2) e la diuresi (acqua
e urea). Ci sono anche piccole perdite di energia nelle feci (cibi non digeriti, cellule derivate dalla desquamazione delle pareti delle mucose, bat

40
Stato e valutazione del metabolismo nell’obesità
teri) e nella miscellanea dei tessuti demoliti (capelli persi, cute, flusso mestruale ecc.) L’introito dei nutrienti deve bilanciare queste perdite secondo i concetti espressi in figura 1 affinché i depositi di energia e la massa corporea rimangano costanti. È altrettanto noto che i principali depositi di energia del corpo sono rappresentati da grassi e proteine; l’altra piccola componente è rappresentata dal glicogeno, generalmente ignorata negli studi perchè poco significativa dal punto di vista calorico e comunque difficilmente misurabile (vedi figura a pagina 3). È difficile stabilire il ruolo effettivo di ciascun nutriente nella formazione dei depositi corporei in quanto ognuno mostra interazioni con l’altro: gli amminoacidi possono essere convertiti a glucosio o a acidi grassi; il glucosio può essere convertito a grasso. Il costo metabolico per convertire il glucosio a glicogeno è pari al 5% del contenuto energetico del glucosio, mentre sale al 24% quando viene convertito a grasso. L’ossidazione dei substrati è dettata dalla necessità di produrre ATP (incremento del rapporto ATP/ADP) e quando questa necessità viene soddisfatta essa viene inibita, a partire dall’inibizione enzimatica della glicolisi, della piruvato deidrogenasi sino alla completa inibizione dell ossidazione degli acidi grassi. In sintesi, la velocità di ossidazione dei macronutrienti per produrre ATP è regolata dalla capacità di riutilizzo del medesimo da parte dell’organismo. A eccezione del tessuto adiposo bruno, nel quale l’ossidazione e la sintesi di ATP sono disaccoppiati, ogni surplus di substrato energetico che non implica una fosforilazione viene depositato. La composizione dei substrati ossidati durante il giorno varia considerevolmente, ma in modo tale da minimizzare la variazione del contenuto proteico e di glicogeno del corpo. Evidentemente l’evoluzione ha preferito sviluppare meccanismi metabolici ed endocrini che agissero sulla regolazione dell’ossidazione dell’intake di proteine e carboidrati, piuttosto che incidere sulle riserve di grasso e sul bilancio energetico. Ciò non deve sorprendere considerando l’importanza delle proteine per le funzioni vitali e del glucosio per il cervello; ciò accade nonostante la riserva corporea di glicogeno sia piccola, ovvero equivalente all’introito di carboidrati usualmente presenti nella nostra dieta quotidiana. È chiaro che la riserva di glicogeno è 50-200 volte più piccola di quella del grasso, e inoltre non rappresenta un’entità distinta e con la sola peculiarità energogenica - ogni grammo di glicogeno s’accompagna con 4 g di acqua, ad esempio - per cui l’organismo trova più facile bilanciare il fabbisogno energetico regolando l’accumulo o la perdita di grasso corporeo. Questa facilità di deposito delle riserve di grasso, unitamente alla relativa difficoltà di mobilizzazione, “urta” la visione di coloro che vedono nella distrettualità adiposa il nemico principale da combattere (visione estetica del corpo), ma certamente costituisce il principale meccanismo

41
Dieta e metabolismo
di adattamento dell’organismo quando sottoposto allo stress di un bilancio energetico negativo (visione salutistica del corpo). È stato più volte rimarcato il fallimento dei programmi di rapido dimagramento, pro-posti nei vari Paesi Occidentali, evidenziando che anche se tali programmi erano volti alla riduzione del grasso percentuale, essi inducevano un decremento ponderale prevalentemente riposto sulla massa magra e un incremento del rischio patologico associato al dimagramento stesso. Evidentemente il tessuto adiposo, oltre che costituire la riserva energetica principale, svolge altre funzioni essenziali nell’organismo regolazione ormonale per esempio - che scaturiscono in una relativa difficoltà di mobilizzazione e utilizzo del medesimo. In sintesi, le variazioni dell’utilizzazione dei substrati durante la restrizione energetica dieta ipocalorica, per esempio - seguono questa scala di priorità per: ■ mantenere la produzione di glucosio da glicolisi e gluconeogenesi
per le necessità tessutali, in particolare per il cervello ■ ridurre al minimo la demolizione delle proteine essenziali del corpo ■ massimizzare infine, la lipolisi e l’ossidazione del grasso allo scopo di
coprire le necessità energetiche, per quanto possibile.
Sorgono spontanee alcune riflessioni di tipo antropologico, che mettono in evidenza la capacità di adattamento metabolico degli ominidi o di alcune popolazioni tuttora viventi, che di fronte alla scarsità di fonti alimentari per lunghi periodi, devono sopravvivere, garantendosi le principali funzioni dell’organismo e bilanciando le necessità energetiche con le riserve di grasso localizzate in precisi distretti corporei. L’ ossidazione dei differenti substrati sembra assumere una precisa gerarchia:
Etanolo > Aminoacidi > Carboidrati > Grassi
Va fatto un inciso, riguardo all’etanolo, che non può essere considerato un macronutriente alla stregua degli altri sopra indicati, ne tantomeno si deposita nel corpo. Da un punto di vista metabolico, il corpo umano sembra una macchina che si muove continuamente (TEE) con la giusta dose di carburante (i macronutrienti), ma se viene alimentata in eccesso è condizionata a bruciare preferenzialmente solo quella parte che si deposita meno facilmente: etanolo, amminoacidi, carboidrati piuttosto che i trigliceridi a lunga catena. Diversi ormoni, insulina, glucagone, catecolamine giocano un ruolo fondamentale nella regolazione dei processi ossidativi e nel bilancio energetico. Le riserve di carboidrati non possono incrementare indefinitamente mentre in teoria la riserva adiposa è senza limite. Inoltre, il pool degli amminoacidi liberi nel sangue regola la dimensione del pool pro

42
Stato e valutazione del metabolismo nell’obesità
teico nel corpo e bilancia l’eccedenza o attraverso la degradazione per via renale o attraverso la conversione. La conseguenza di questa gerarchia ossidativa è che ogni macronutriente in eccesso tende a convertirsi in grasso corporeo. Un surplus calorico derivato da un pasto aggiuntivo, composto esclusivamente di grassi, porta a un incremento delle riserve adipose con un efficienza metabolica del 99%. Pensate al classico gelato con abbondante panna dopo una lauta cena serale; oppure alle piccole porzioni di formaggio o di dolce con crema, porzioni consumate sempre al termine del pasto come premianti stuzzichini in risposta alla giornata di stress, più che elementi sazianti! Ebbene questi piccoli errori nutrizionali hanno nel breve termine un effetto “infausto”: il costo metabolico per il deposito della componente lipidica è praticamente nullo. Ma l’eccedenza in carboidrati? Secondo le osservazioni di Acheson KJ e Flatt JP , una grande quantità di carboidrati (480 g), fornita a soggetti nella norma e dopo un digiuno notturno, induce l’ossidazione e il ripristino della scorta glicogenica, mentre solo in minima parte si converte in trigliceridi di deposito (tabella 1). Questo implicherebbe che in un soggetto con bilancio energetico negativo, (ad esempio durante un regime dimagrante), una dieta con un alto rapporto di carboidrati rispetto ai grassi non porta all’obesità. Pensate alla fobia dei tanti pazienti in dieta ipocalorica nei confronti di pasta, pane e altri carboidrati. Pensate ai “sensi di colpa” di coloro che vedono in un piatto di spaghetti la massima trasgressione del loro perenne regime di dieta. In aggiunta è stato più volte dimostrato che il pattern dei depositi adiposi riflette la qualità della dieta assunta, mostrando una stretta correlazione con la percentuale lipidica della dieta stessa piuttosto che con i processi di biosintesi. Se tali osservazioni sono valide nei regimi dimagranti, esse vengono tuttavia, a decadere in altre circostanze. Ad esempio nei soggetti malnutriti, alimentati in nutrizione parenterale totale, anche il pasto composto esclusivamente di glucidi può indurre una lipogenesi de novo. Se poi iperalimentiamo (Jequier et Al) sempre con car-
TABELLA 1
A sei soggetti nella norma sono stati forniti 478 g di CHO dopo digiuno notturno. Lo studio effettuato con calorimetria indiretta si è
protratto per 10 ore dopo l’ingestione del pasto. I risultati sono i seguenti:
■ 29 g di proteine ossidate
■ 133 g di CHO ossidati
■ Perciò 346 g di CHO sono stati immagazzinati come glicogeno
■ 17 g di grasso ossidati
La tabella mostra i risultati relativi all’introduzione di un pasto abbondante composto esclusivamente di carboidrati.
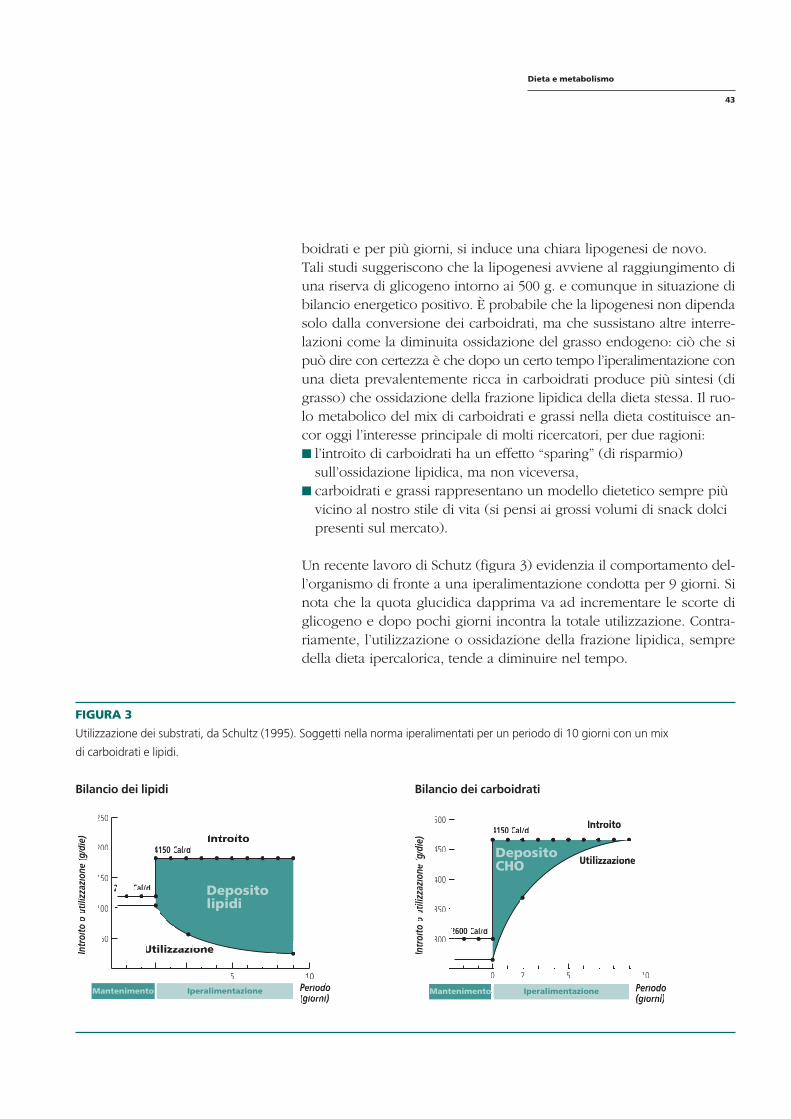
0 20
43
250
Dieta e metabolismo
boidrati e per più giorni, si induce una chiara lipogenesi de novo. Tali studi suggeriscono che la lipogenesi avviene al raggiungimento di una riserva di glicogeno intorno ai 500 g. e comunque in situazione di bilancio energetico positivo. È probabile che la lipogenesi non dipenda solo dalla conversione dei carboidrati, ma che sussistano altre interrelazioni come la diminuita ossidazione del grasso endogeno: ciò che si può dire con certezza è che dopo un certo tempo l’iperalimentazione con una dieta prevalentemente ricca in carboidrati produce più sintesi (di grasso) che ossidazione della frazione lipidica della dieta stessa. Il ruolo metabolico del mix di carboidrati e grassi nella dieta costituisce ancor oggi l’interesse principale di molti ricercatori, per due ragioni: ■ l’introito di carboidrati ha un effetto “sparing” (di risparmio)
sull’ossidazione lipidica, ma non viceversa, ■ carboidrati e grassi rappresentano un modello dietetico sempre più
vicino al nostro stile di vita (si pensi ai grossi volumi di snack dolci presenti sul mercato).
Un recente lavoro di Schutz (figura 3) evidenzia il comportamento del-l’organismo di fronte a una iperalimentazione condotta per 9 giorni. Si nota che la quota glucidica dapprima va ad incrementare le scorte di glicogeno e dopo pochi giorni incontra la totale utilizzazione. Contrariamente, l’utilizzazione o ossidazione della frazione lipidica, sempre della dieta ipercalorica, tende a diminuire nel tempo.
FIGURA 3
Utilizzazione dei substrati, da Schultz (1995). Soggetti nella norma iperalimentati per un periodo di 10 giorni con un mix
di carboidrati e lipidi.
Bilancio dei lipidi Bilancio dei carboidrati
500
105
Introito
Utilizzazione
Deposito lipidi
4150 Cal/d
2600 Cal/d
Intr
oito
o u
tiliz
zazi
one
(g/d
ie)
Introito
UtilizzazioneDeposito CHO
4150 Cal/d
2600 Cal/d
Intr
oito
o u
tiliz
zazi
one
(g/d
ie)
450
400
350
300
0 2 5 10
Mantenimento Iperalimentazione Periodo Mantenimento Iperalimentazione Periodo (giorni) (giorni)
50
100
150
200

44
Stato e valutazione del metabolismo nell’obesità
Il meccanismo che spiega tale effetto potrebbe essere il seguente: ■ diminuisce l’ossidazione lipidica poichè diminuisce la lipolisi, attività
mediata dall’incremento dei livelli di insulina data la forte presenza di glucidi,
■ la quota lipidica della dieta viene integralmente convertita in trigliceridi di deposito.
Nel breve termine sembra che l’organismo non abbia capacità di regolare il bilancio del grasso; per esempio è stato dimostrato, che dopo 9 ore, un individuo in normopeso alimentato con un supplemento di due pasti, di cui uno con 50 g di lipidi in più, immagazzinava solamente il pasto lipidico, in quantità di grasso pari esattamente ai 50 g aggiunti. Una simile osservazione veniva confermata anche dopo un periodo di 1-2 giorni con un paritetico supplemento di lipidi. Il gruppo di Frayn K.N. in un’altro test condotto su soggetti normali, in cui si comparavano due pasti - uno con 80 g di zuccheri, l’altro con 80 g di zuccheri più 80 g di lipidi- trovò che nonostante l’incremento dell’ossidazione del substrato lipidico, ben 70 g di grasso venivano depositati con un risparmio sul l’ossidazione dei carboidrati pari a solo 20 g. Occorre rimarcare che è il supplemento di grasso, cioè quello addizionato alla dieta supposta isoenergetica, che viene rapidamente immagazzinato. La difficoltà di trarre conclusioni definitive da questi studi è legata al fatto che i pasti test non sono tra loro isoenergetici: si può potrebbe unicamente arguire che l’organismo risponde al surplus calorico da grasso, depositando il grasso stesso per il riequilibrio energetico. Negli esperimenti a breve termine vi è, quindi, una stretta relazione tra il bilancio del grasso e quello energetico; al contrario, numerosi lavori non dimostrano alcuna relazione tra il bilancio proteico - glucidico e quello energetico. Nella tabella 2 vengono riassunte le principali caratteristiche dei macronutrienti e il loro ruolo nel bilancio metabolico. è interessante notare anche la diversità del comportamento dei macro-nutrienti nella regolazione dell’intake e dell’appetito. Secondo Schutz (93) si può ipotizzare un modello di calcolo del guadagno ponderale su un soggetto iperalimentato. Ritornando a quanto espresso inizialmente, un eccesso di 100 cal da lipidi, per un individuo di 60 kg, comporterebbe un incremento di 7,5 kg in 6 anni, di cui 2/3 (5,1 kg) sono costituiti da massa grassa depositata. Sulla base di passate risul lipidica pari a 2 g al giorno per ogni kg di grasso depositato, per cui alla fine del periodo, si ossiderebbero 10 g pari a 90 cal. Solo dopo 6 anni l’ossidazione lipidica bilancia l’eccesso alimentare: il peso, composto in prevalenza di massa grassa, nel frattempo continua ad aumentare. In genere, l’incremento della massa adiposa è correlata al metabolismo post-assorbitivo, che riflette un aumentato effetto sui NEFA (acidi gras
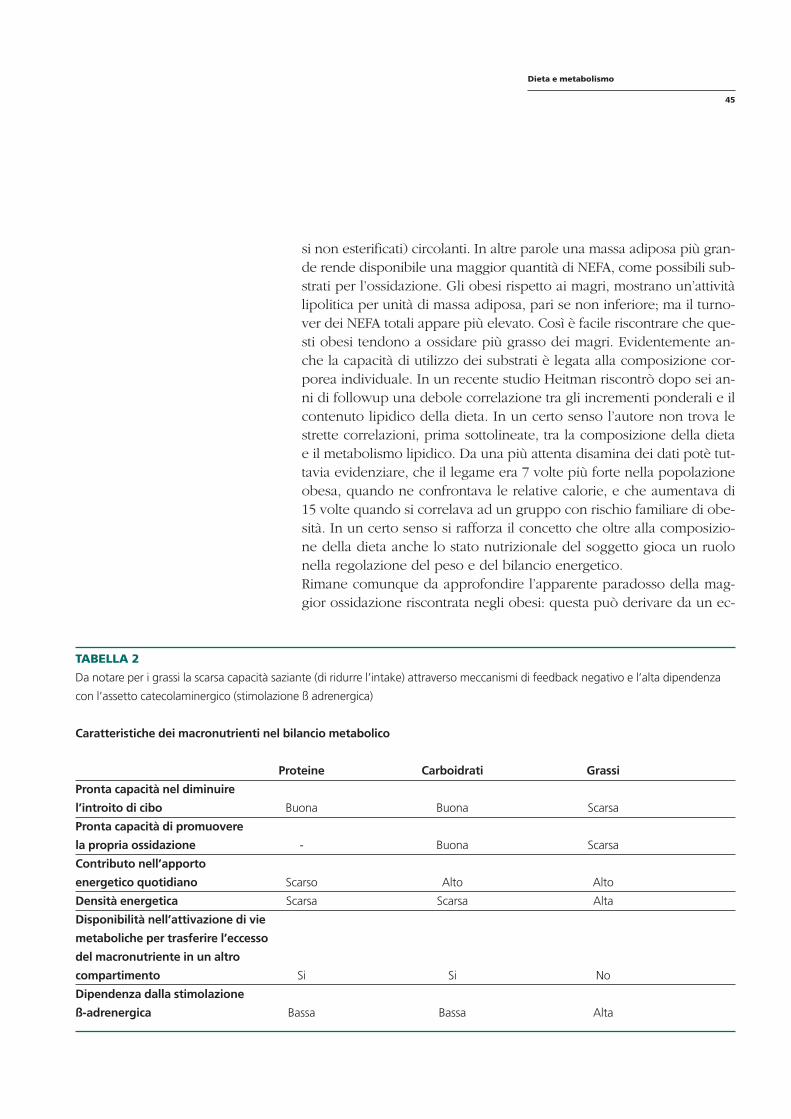
45
Dieta e metabolismo
si non esterificati) circolanti. In altre parole una massa adiposa più grande rende disponibile una maggior quantità di NEFA, come possibili substrati per l’ossidazione. Gli obesi rispetto ai magri, mostrano un’attività lipolitica per unità di massa adiposa, pari se non inferiore; ma il turnover dei NEFA totali appare più elevato. Così è facile riscontrare che questi obesi tendono a ossidare più grasso dei magri. Evidentemente anche la capacità di utilizzo dei substrati è legata alla composizione corporea individuale. In un recente studio Heitman riscontrò dopo sei anni di followup una debole correlazione tra gli incrementi ponderali e il contenuto lipidico della dieta. In un certo senso l’autore non trova le strette correlazioni, prima sottolineate, tra la composizione della dieta e il metabolismo lipidico. Da una più attenta disamina dei dati potè tuttavia evidenziare, che il legame era 7 volte più forte nella popolazione obesa, quando ne confrontava le relative calorie, e che aumentava di 15 volte quando si correlava ad un gruppo con rischio familiare di obesità. In un certo senso si rafforza il concetto che oltre alla composizione della dieta anche lo stato nutrizionale del soggetto gioca un ruolo nella regolazione del peso e del bilancio energetico. Rimane comunque da approfondire l’apparente paradosso della maggior ossidazione riscontrata negli obesi: questa può derivare da un ec-
TABELLA 2
Da notare per i grassi la scarsa capacità saziante (di ridurre l’intake) attraverso meccanismi di feedback negativo e l’alta dipendenza
con l’assetto catecolaminergico (stimolazione ß adrenergica)
Caratteristiche dei macronutrienti nel bilancio metabolico
Proteine Carboidrati Grassi
Pronta capacità nel diminuire
l’introito di cibo Buona Buona Scarsa
Pronta capacità di promuovere
la propria ossidazione - Buona Scarsa
Contributo nell’apporto
energetico quotidiano Scarso Alto Alto
Densità energetica Scarsa Scarsa Alta
Disponibilità nell’attivazione di vie
metaboliche per trasferire l’eccesso
del macronutriente in un altro
compartimento Si Si No
Dipendenza dalla stimolazione
ß-adrenergica Bassa Bassa Alta

46
Stato e valutazione del metabolismo nell’obesità
cesso di NEFA circolanti o più semplicemente può essere un ipotesi valida solo per gli individui che hanno un’obesità datata e stabile e non per quelli che la stanno sviluppando: a significare, che per i primi, il processo ossidativo viene facilmente stimolato per contenere eventuali incrementi di peso. Alcuni lavori sembrano confluire su queste ipotesi, quando evidenziano che chi ha una predisposizione all’obesità, mostra, di fronte a una dieta iperlipidica, una riduzione della capacità ossidativa del substrato lipidico. Di converso, l’insulino resistenza, facilitando l’ossidazione lipidica rispetto a quella glucidica, diminuisce il rischio dell’incremento ponderale. Come la massa adiposa e l’insulino-resistenza aumentano, l’ossidazione lipidica cresce sino ad equilibrare l’introito di grasso con la dieta. Il flusso degli acidi grassi, da e per il tessuto adiposo, è regolato da numerosi fattori, ma in particolare risulta strettamente condizionato da quelli associati all’eccedenza energetica - insulina, glucosio, triacilglicerolo plasmatici soprattutto che favorirebbero l’uptake e l’esterificazione degli acidi grassi piuttosto che il rilascio. Nel lungo termine, nel corso della vita ad esempio, una simile facilità all’immagazzinamento del grasso dovrebbe indurre ad un illimitata riserva corporea del medesimo. Questo ha portato a supporre l’esistenza di un meccanismo di regolazione dei depositi adiposi (teoria lipostatica), visto che la maggior parte della popolazione comunque presenta un peso relativamente stabile con una alta precisione (1%) nella regolazione del bilancio della massa lipidica corporea. La teoria lipostatica suggerisce una interrelazione tra l’introito di grasso esogeno e le riserve di grasso corporeo, cioè un meccanismo di feedback che blocca l’introito di grasso di fronte ad un alta percentuale del medesimo nel corpo. A supporto di tale teoria si sono sviluppati numerosi lavori, a partire da quelli curati da Kennedy nel 1953, che poneva l’attenzione sui meccanismi di regolazione dell’intake e della sazietà per via ipotalamica, sino a giungere ai più recenti di Zhang legati alla scoperta del gene ob. Il prodotto del gene, un piccolo peptide chiamato leptina, prodotto dal tessuto adiposo e scoperto per la prima volta nei ratti, avrebbe un suo omologo umano capace di regolare con un meccanismo di feedback l’appettito. Attualmente i numerosi studi sperimentali della leptina sull’uomo, in particolare i suoi possibili effetti sulla regolazione dell’intake dei pazienti obesi, troverebbero il loro razionale d’uso nella supposta esistenza di una alterazione del meccanismo regolatorio lipidico. Il condizionale in questo ambito è d’obbligo, in quanto gli studi sull’uomo si stanno, solo oggi, sviluppando e in base anche ad alcune risultanze che portano alla constatazione di effetti diversi rispetto al topo. È possibile tuttavia un’altra ipotesi: il bilancio della massa lipidica corporea potrebbe ri

47
Dieta e metabolismo
L’ADATTAMENTO METABOLICO: SCOPI E CONSEGUENZE
flettere nel lungo termine lo stretto legame con il bilancio energetico (Teoria Energostatica). Se l’organismo ha in sè il codice di mantenere il bilancio energetico, poiché il ruolo del grasso corporeo è quello di maggior riserva energetica per il lungo termine, allora anche il bilancio del grasso deve essere mantenuto nel corso della vita In questo caso si pone l’accento più al surplus energetico che alla qualità o composizione della dieta; questo tuttavia non esclude che se il surplus è rappresentato da grassi lo sviluppo di obesità viene favorito. In effetti, un surplus energetico non condiziona indistintamente l’EI, con un paritetico feedback negativo su ogni singolo macronutriente.
Si è già accennato al “punto di vista evolutivo” sottolineando che l’organismo, sin dagli albori della specie attuale, presentava selezionati meccanismi di adattamento energetico per proteggere la massa magra dal la carenza periodica di cibo e dalla fame ricorrente. Il principio di adattamento metabolico è quindi quello di attivare un numero di meccanismi di risparmio energetico per proteggere le funzioni vitali finché è possibile. In base a tale principio, il corpo mostra “resistenza” ai regimi ipocalorici, mentre all’opposto, mostra una grande “sensibilità recettiva” nei periodi di abbondanza del cibo. Il problema è che resistenza e sensibilità si traducono spesso nella capacità di immagazzinare rapidamente tessuto grasso, a salvaguardia della massa magra e delle sue componenti. Coerentemente, l’organismo sottoposto allo stress ipocalorico persegue i seguenti obbiettivi: ■ mantenere la glicolisi e la gluconeogenesi per le necessità tessutali e
degli organi ■ ridurre al minimo la demolizione proteica ■ attivare l’ossidazione del grasso corporeo solo quando la richiesta
energetica diviene protratta e, come tale, può andare a discapito delle prime due funzioni.
Oggi, la popolazione attua arbitrariamente i periodi di restrizione energetica per raggiungere livelli di peso corporeo all’interno di margini ideali, quanto inaccettabili. L’autogestione nei programmi di dimagramento, cioè l’assenza del medico, vede la scelta di modelli ipocalorici VLCD (very low caloric diet <1000 Cal) causa nel tempo di malnutrizione, rischio cardiovascolare e recupero ponderale. Gli effetti conseguenti ai rapidi e drastici dimagramenti mostrano, nella loro evidenza clinica, la deplezione delle componenti idrica, proteica e

48
Stato e valutazione del metabolismo nell’obesità
LA RISPOSTA METABOLICA ALLA RIDUZIONE DELL’APPORTO ENERGETICO
minerale della massa magra e la scarsa risposta sulla massa grassa. È la conferma della teoria che vede nell’impulso prioritario di tesaurizzare le scorte energetiche la garanzia che solo un meccanismo “energy-saving” può mantenere le funzioni vitali corporee. In prima analisi, sorprende la perdita immediata di massa magra quasi ad informarci che l’organismo sotto lo stress di un bilancio energetico negativo voglia proprio ridurre le sue funzioni vitali; ma come vedremo più avanti vengono attivati diversi meccanismi a conferma del principio energostatico e a salvaguardia delle funzioni vitali. Gluconeogenesi e glicolisi, sintesi e ossidazione proteica, rappresentano in questa logica i meccanismi iniziali per fornire rispettivamente energia (glucosio) al sistema nervoso centrale, agli organi, e aminoacidi per mantenere costante il pool di AA liberi. Questi meccanismi sono rapidi ed efficienti. Ma di fronte al protrarsi del bilancio energetico negativo, la mobilizzazione e l’ossidazione delle riserve lipidiche corporee non sono altrettanto rapide e la riduzione del fabbisogno basale (BMR) appare come la risposta più immediata: un numero di meccanismi che almeno parzialmente potrebbero essere responsabili dell’insuccesso a lungo termine nel dimagramento o nel mantenimento del peso. Il quesito dei prescrittori di diete ipocaloriche riguarda il livello di calorie che può garantire il maggior decremento ponderale, mentre sarebbe più opportuno porre l’attenzione su come limitare la diminuizione della massa magra e il conseguente adattamento metabolico.
La letteratura ci offre numerosi spunti riguardo alla risposta metabolica a partire dagli esempi più drastici nelle loro proposizioni. Ad esempio, nel Minnesota Study del 1940, Keys studiò per 24 settimane gli effetti le del semidigiuno su trentadue volontari magri con una dieta di 650 calorie circa. Durante questo periodo persero il 23% del loro peso corporeo iniziale. Contemporaneamente il BMR diminuiva del 36%, specialmente la massa muscolare che diminuiva di circa il 40%, mentre gli organi rimanevano relativamente immutati. In particolare la massa magra corporea (LBM) diminuiva del 26%. La diminuzione di BMR per unità di LBM evidenziava un 14% in meno, a conferma dei meccanismi di adattamento. Questa risposta potrebbe rappresentare un’aumentata “efficienza metabolica della rimanente LBM”. Grande et al. consideravano la diminuzione nell’attività metabolica della LBM come il principale fattore responsabile della BMR ridotto, ma le loro analisi provenivano da studi sperimentali a breve termine (< 4 settimane). Al contrario Keys ed altri nel loro studio giunsero alla conclusione che circa due terzi della diminuzione di BMR era originata dalla riduzione di LBM.

49
Dieta e metabolismo
Queste differenze stanno a significare che forse solo nel lungo termine intervengono i meccanismi di adattamento. Nell’esperimento del Minnesota non era stato valutato il dispendio energetico totale (TEE) e quindi non si poteva discriminare tra le eventuali alterazioni delle componenti relative all’attività fisica. Ma le misurazioni indirette di altri fattori, come la stanchezza e l’apatia, hanno chiaramente mostrato che questa componente del dispendio non era implicata. Recentemente Prentice e colleghi hanno pubblicato dati di metaanalisi riguardanti 29 studi sulle risposte metaboliche al dimagramento. Questi studi, contrariamente allo studio nel Minnesota, erano principalmente rivolti ai soggetti obesi, e quindi possono essere un utile riferimento. È d’obbligo domandarci se in tali circostanze, quando c’è un’abbondanza di massa grassa, la riduzione metabolica sia la stessa. Persino nelle più drastiche VLCD, era chiaramente evidente che la diminuzione nella componente metabolica raggiungeva un plateau oscillante tra il 20% ed il 25%. La differente risposta rispetto al Minnesota Study -un minor calo di BMR - può essere spiegata dalla massa grassa iniziale più elevata: per bilanciare la restrizione energetica i soggetti obesi si “affidano” alle loro scorte di grasso e perdono una percentuale inferiore di tessuto attivo, magro, rispetto ai soggetti normopeso. Da questa metanalisi si apprende tuttavia, che l’effetto di massimo adattamento del BMR per unità di LBM è minore del 15% e quindi che si ha comunque un adattamento metabolico. Viene spontanea l’osservazione che il regime ipocalorico determina il meccanismo di adattamento sia sui magri che sugli obesi e che la riduzione dell’apporto energetico può comportare una diminuizione dell’attività metabolica non continua: si raggiunge infatti un plateau oltre il quale si potrebbe ipotizzare l’assenza di meccanismi che salvaguardano le funzioni vitali. Per quanto tempo quindi si deve modulare la restrizione calorica? E a sua volta come si modula la risposta metabolica? James e Shetty hanno mostrato che l’adattamento metabolico (per esempio un efficienza metabolica aumentata) è visibile nelle prime due settimane di riduzione energetica. Ma è importante soprattutto lo studio di De Boer che vede già nella prima fase di restrizione una rapida riduzione del BMR: dopo 24 ore con una dieta di 1000 cal/die il TEE diminuisce del 5%. Questo meccanismo , indipendente dalle variazioni nella composizione corporea, era inevitabile. Alla fine dell’ottava settimana del periodo di dieta, e dopo un giorno di rialimentazione con dieta di mantenimento, si conseguiva un recupero del TEE del 6%. L’autore attribuisce alla diminuita termogenesi (DIT), conseguente a un minor apporto di cibo, il pronto calo metabolico. La componente DIT è parzialmente mediata dal SNS (Sistema Nervoso Simpatico). La seconda risposta di adattamento è relativa alla diminuzione nel peso corporeo nella sua LBM. Negli studi di De Boer

50
Stato e valutazione del metabolismo nell’obesità
questa componente diminuiva del 5% al mese. Sembra quindi ragionevole affermare che la riduzione in BMR nel periodo di dieta ipocalorica avviene in due differenti fasi: ■ la fase iniziale, attribuibile all’aumento dell’efficienza metabolica
della restante LBM, e attribuibile alla diminuita termogenesi del pasto ■ la fase successiva, principalmente giustificata dall’ulteriore calo nel
BMR dalla perdita di LBM.
Tale disamina fa sorgere alcune riflessioni sui modelli di dieta che vengono prescritti. La somministrazione del regime ipocalorico, come modello da adottare sin dal primo giorno, crea un immediato adattamento (-5%) e quindi penalizza ulteriormente l’efficacia della dieta. Eppure la maggior parte dei medici, cosi’ vincolata dalla promessa di un calo ponderale importante, al quale purtroppo lega il successo della sua prestazione, tende a partire con la proposta calorica più restrittiva. Occorre partire con soluzioni graduali, ma questo impone una capacità da parte del medico di “legare” il paziente con altri metodi, indipendentemente o contrariamente all’assillante richiesta di perdere subito peso. Piccole modifiche relative alla distribuzione e alla qualità degli alimenti (tabella 2), un supporto di educazione e terapia comportamentale, appaiono le soluzioni che potrebbero essere attivate in alternativa all’immediata prescrizione di un regime restrittivo. Ma tutto questo apre il problema delle reali competenze del medico nel-l’area dell’educazione e della terapia comportamentale. In anni recenti queste aree sono state ampiamente sviluppate senza tuttavia determinare un approccio pratico e applicativo, ad eccezione delle proposte metodologiche di pochi esperti, quali: Brownell, Kirshenbaum, Melchionda, Dalle Grave per citarne alcuni. Poichè la dieta viene usualmente consigliata per periodi protratti sarebbe opportuno valutare quale livello calorico produce il minimo adattamento. Prentice, combinando i dati di 22 studi con una durata di 4-6 settimane, trovò con diete inferiori a 850 cal giornaliere una diminuizione del 15% in BMR rispetto al 5% da diete di circa 1250 cal. Essi conclusero che c’è una soglia a circa 1250 cal., al di sotto della quale interviene una difesa fisiologica più forte contro la perdita di peso. La perdita di peso era circa del 2% a settimana per le diete con un apporto energetico molto basso in rapporto all’1.3% per le diete con una riduzione moderata di energia. La differenza tuttavia, indica una perdita più alta di LBM e non di massa grassa! La terza osservazione è che i regimi ipocalorici protratti devono essere superiori alle 1250 cal. Un tale suggerimento, ovviamente, non rappresenta la regola, poichè la prescrizione dovrebbe sortire sempre dalla disamina del reale stato nutrizionale del paziente in esame. Il paziente ad esempio potrebbe aver subito pregresse esperienze di fluttuazione del

51
Dieta e metabolismo
peso, oppure potrebbe avere una LBM significativa, o un’attività fisica attuale al di sopra dei livelli precodificati. Occorre com’è noto partire dalla storia clinica, dalla disamina del consumo alimentare, dall’analisi della composizione corporea, dalla misura del BMR, sino alla valutazione dei parametri laboratoristici. Generalmente una perdita di massa corporea, con un rapporto di 1:3 di LBM rispetto alla massa grassa, è accettato come normale per gli individui obesi. La maggior perdita di LBM durante una dieta drastica avviene nel breve periodo. Specialmente durante le prime settimane di una VLCD, la perdita di LBM è più elevata a causa del forte gap energetico iniziale e del lento avvio, sempre iniziale, della lipolisi massimale e dell’ossidazione del grasso. I dati di numerosi studi riguardanti la VLCD ed effettuati per un periodo superiore ai tre mesi non hanno comunque rivelato una perdita di LBM più grande del 25%. Questo conferma l’ipotesi, che se la durata della dieta è sufficientemente lunga, le modificazioni nel primo periodo in cui si perde più peso, scompariranno nel processo totale di adattamento, sino a mantenere una risposta metabolica costante: il paziente perde a fatica il peso poichè diminuisce la perdita di massa magra e si coinvolge sempre di più la massa grassa.

Gestire il paziente
in sovrappeso:
una proposta concreta
e completa
per la diagnosi,
la terapia e il controllo.
30 anni fa Dietosystem lanciava il suo primo sistema computerizzato di dietoterapia. Nasceva così un approccio nuovo al trattamento del sovrappeso e delle patologie correlate.
Da allora questo strumento è stato costantemente perfezionato e aggiornato, e tutt’oggi è leader nel settore. Da allora Dietosystem ha sviluppato il suo approccio originale, affiancando alla dietoterapia una metodologia sempre più vasta e rappresentativa di un insieme unico di tecnologie e presidi mirati all’area del sovrappeso della obesità e delle patologie associate. Logico® è l’espressione concreta del nuovo sistema esperto ove la metodologia gestionale del paziente in sovrappeso e obeso segue la traccia di un protocollo scientifico ideale. Grazie a Logico®, Dietosystem traduce il protocollo ideale in una serie di step diagnostici e terapeutici nei più diversi set clinici: dal reparto ospedaliero al poliambulatorio e allo studio medico del singolo professionista.
In campo diagnostico, la metodologia Dietosystem offre tecniche non invasive, con un eccellente rapporto tra costi e benefici, nelle aree della composizione corporea, del food intake e comportamento alimentare, del metabolismo, del microcircolo e della patologia venosa.
In campo terapeutico Dietosystem offre apparecchiature e presidi ad alta tecnologia in grado di trattare le adiposità localizzate, gli stati edemigeni e la patologia del microcircolo venoso. Dove non arriva lo strumento le nuove linee di supplementi offrono opzioni terapeutiche sistemiche o topiche per una rapida soluzione del quadro clinico.
Con le stesse caratteristiche di affidabilità ed efficacia, Dietosystem ha messo a punto strumenti per elaborare e applicare programmi di attività fisica mirati alla riduzione del sovrappeso e delle sue complicanze. Le soluzioni elaborate da Dietosystem costituiscono quindi un sistema integrato e completo di gestione altamente sofisticato quanto amichevole nell’utilizzo. Un sistema unico a livello internazionale perché traduce le esperienze concrete di 30 anni di rapporto diretto con il medico.
20125 Milano - Viale Monza, 133 Tel. 02 2817 2200 - Fax 02 2817 2299
eMail: [email protected] Web: www.dsmedica.info

&







