

Il glucosio può provenire dalla dieta, in quanto tale, o
derivare dalla demolizione di carboidrati complessi.
Viene metabolizzato per via aerobica o anaerobica, fornendo energia
all’organismo.
E’ l’unica fonte energetica per i globuli rossi (metabolismo anaerobico) e
la principale per il cervello (metabolismo aerobico).
E’ indispensabile per un efficiente catabolismo dei grassi.
La principale fonte di glucosio nella dieta è l’amido.
IL GLUCOSIO
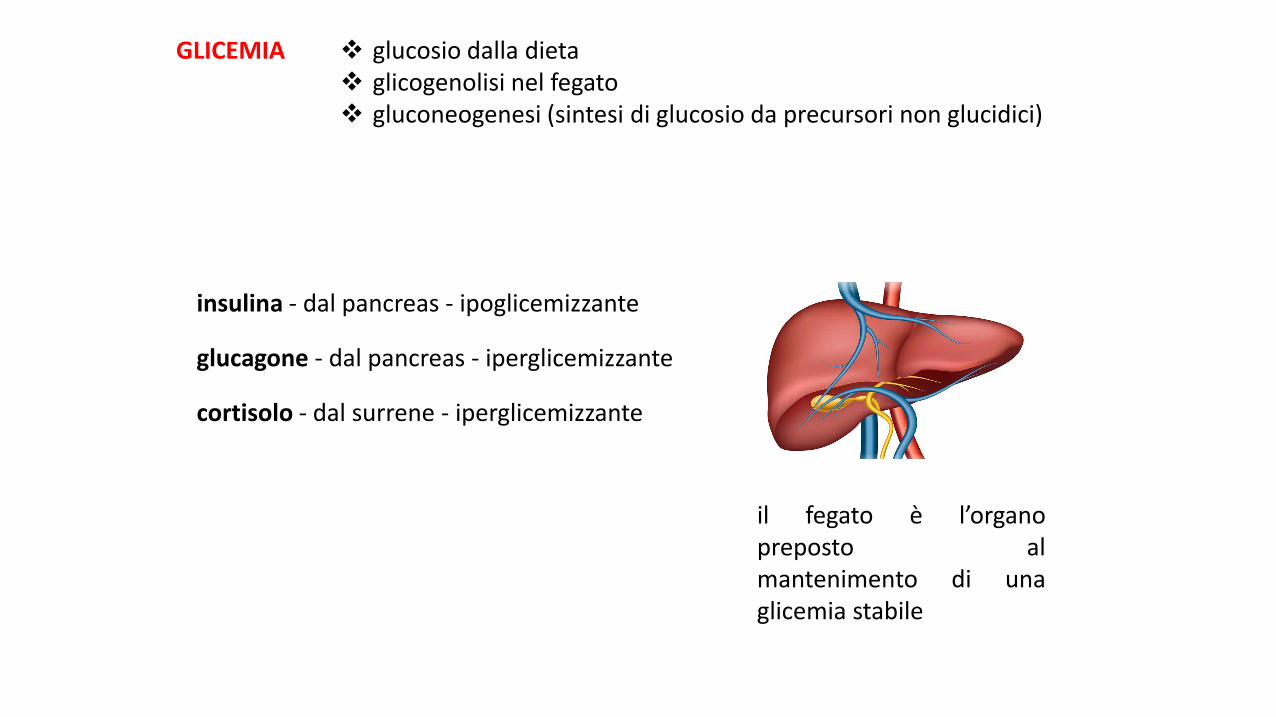
il fegato è l’organopreposto almantenimento di unaglicemia stabile
GLICEMIA glucosio dalla dieta glicogenolisi nel fegato gluconeogenesi (sintesi di glucosio da precursori non glucidici)
insulina - dal pancreas - ipoglicemizzante
glucagone - dal pancreas - iperglicemizzante
cortisolo - dal surrene - iperglicemizzante

3
GLICOLISI È una via metabolica rapida, che può fornireenergia sia in presenza che in assenza di O2.
6 CO2 + 6 H2O
O2
acido lattico
acido piruvico
4
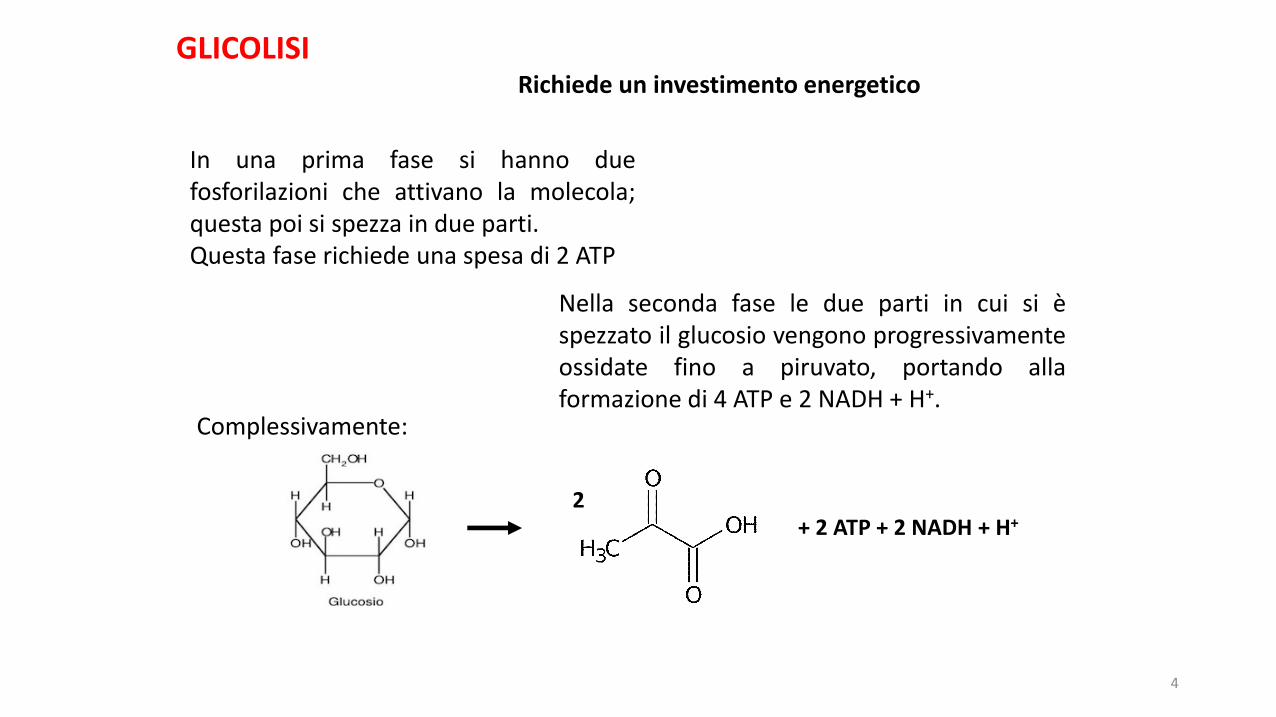
Richiede un investimento energetico
In una prima fase si hanno duefosforilazioni che attivano la molecola;questa poi si spezza in due parti.Questa fase richiede una spesa di 2 ATP
Nella seconda fase le due parti in cui si èspezzato il glucosio vengono progressivamenteossidate fino a piruvato, portando allaformazione di 4 ATP e 2 NADH + H+.
GLICOLISI
Complessivamente:
2+ 2 ATP + 2 NADH + H+
5
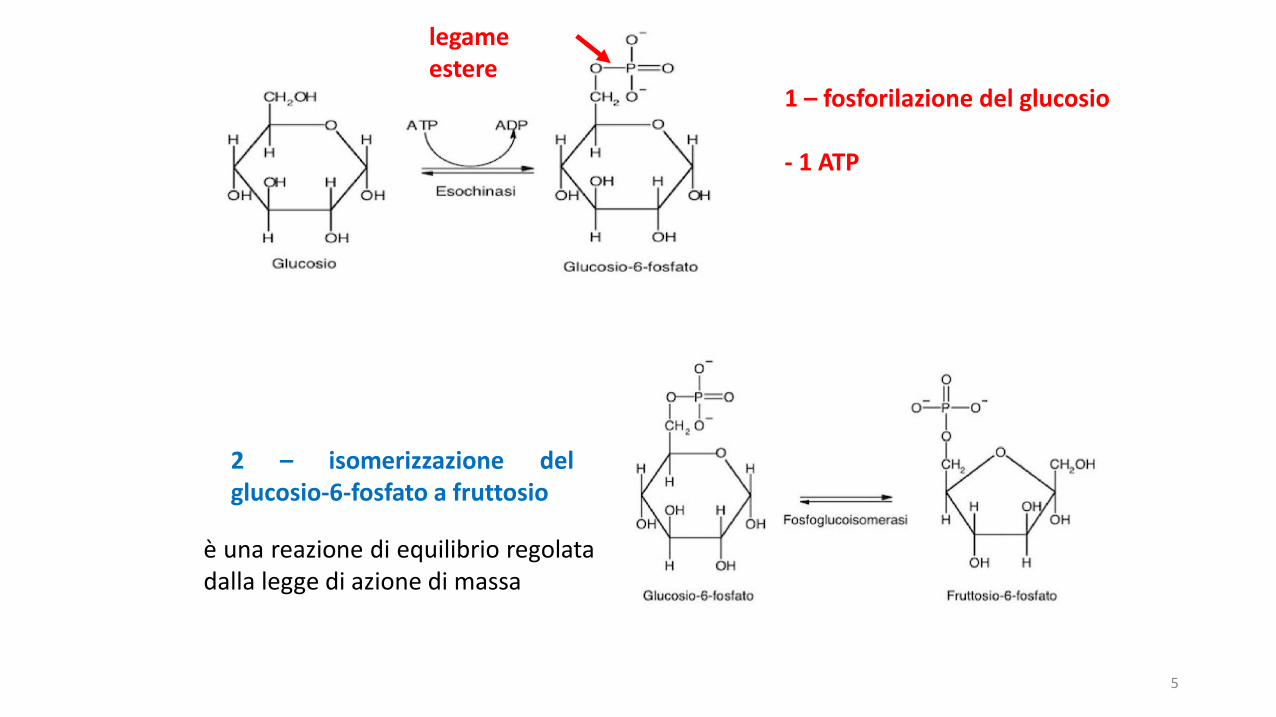
legame estere
1 – fosforilazione del glucosio
- 1 ATP
2 – isomerizzazione delglucosio-6-fosfato a fruttosio
è una reazione di equilibrio regolatadalla legge di azione di massa

6
3 – fosforilazione del fruttosio-6-fosfato
- 1 ATP
La trasformazione è catalizzata dall’enzimafosfofruttochinasi; è stata consumata un’altramolecola di ATP e ora il fruttosio dispone di duegruppi attivati.
fruttosio-6-fosfato fruttosio-1,6-bifosfato

7
4 – scissione del fruttosio-1,6-bifosfato
- 1 ATP
fruttosio-1,6-bifosfato
O-PO32-
CH2
CH2O-PO32-
C O
CHOH
CH OH
CH OH
ora sono disponibili due triosi in forma attivata

i due triosi possono trasformarsi l’uno nell’altro;l’unico che entra nel ciclo metabolico è la gliceraldeide
.
la trasformazione è catalizzata dall’enzima trioso fosfato isomerasi
diidrossiacetonefosfato
gliceraldeide-3-fosfato
da qui in poi, quindi, ogni reazione andrà contata due volte, perchéavverrà su due molecole di gliceraldeide-3-fosfato.
5 – interconversione dei triosi
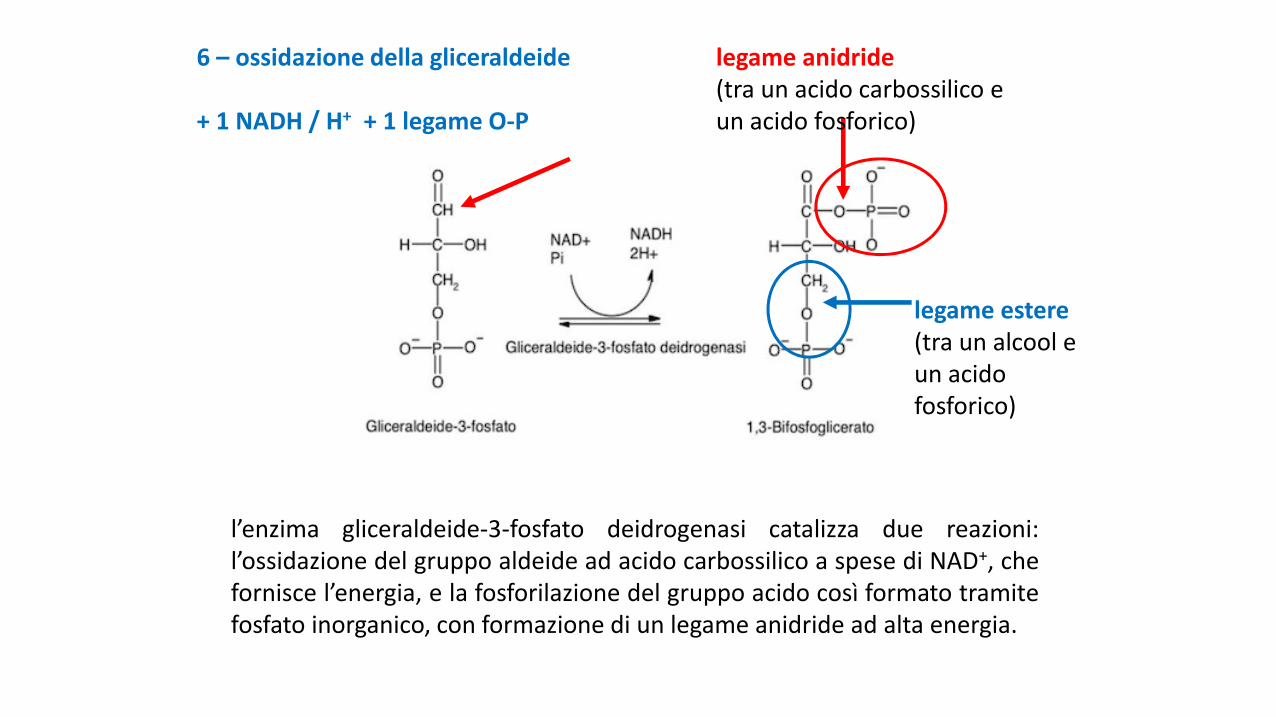
6 – ossidazione della gliceraldeide
+ 1 NADH / H+ + 1 legame O-P
l’enzima gliceraldeide-3-fosfato deidrogenasi catalizza due reazioni:l’ossidazione del gruppo aldeide ad acido carbossilico a spese di NAD+, chefornisce l’energia, e la fosforilazione del gruppo acido così formato tramitefosfato inorganico, con formazione di un legame anidride ad alta energia.
legame anidride(tra un acido carbossilico e un acido fosforico)
legame estere(tra un alcool e un acido fosforico)

7 – produzione di 3-fosfoglicerato
+ 1 ATP
il gruppo fosforico viene trasferito dal legame anidride delbifosfoglicerato all’ATP; il legame estere dell’altro gruppo fosforico nonha sufficiente energia per fosforilare l’ADP.

8 – trasferimento di fosfato

enolasi
9 – produzione di fosfoenolpiruvato (PEP)
+ 1 legame ad alta energia
è una disidratazione: si perde OH- dal C3 e H+ dal C2,con formazione di un doppio legame (eliminazione).
0+1
-1 -2
in seguito alla disidratazione vi è un notevole cambiamento nelladistribuzione dell’energia all’interno della molecola, a seguito delquale il G° di idrolisi del gruppo fosforico passa da -4,2 kcal/molnel fosfoglicerato a – 14,8 kcal/mol nel PEP

10 – produzione di piruvato
+ 1 ATP
Quello che ora è un legame fosforico a maggiorenergia può essere usato per fosforilare l’ADP.
C
CH3
O
CO–
O
Si ottiene acido piruvico, che può poi esseretrasformato in AcetilCoA ed entrare nel ciclo di Krebs,oppure essere utilizzato come precursoredell’amminoacido alanina

BILANCIO DELLA GLICOLISI
Glucosio
Glucosio-6-P
Fruttosio-6-P
Fruttosio-1,6-bis-P
Gliceraldeide-3-P Diidrossiacetone-P
ATP
ATP
fase di investimento energetico
Gliceraldeide-3-P
1,3-bis-P-glicerato
3-P-glicerato
2-P-glicerato
Fosfoenolpiruvato
Piruvato
fase di produzione di energia
ATP
X 2
ATPNADH
Glucosio 2 CH3CO-COOH + 2 ATP + 2 NADH + H+
le tappe indicate da una singolafreccia sono irreversibili (G << 0)
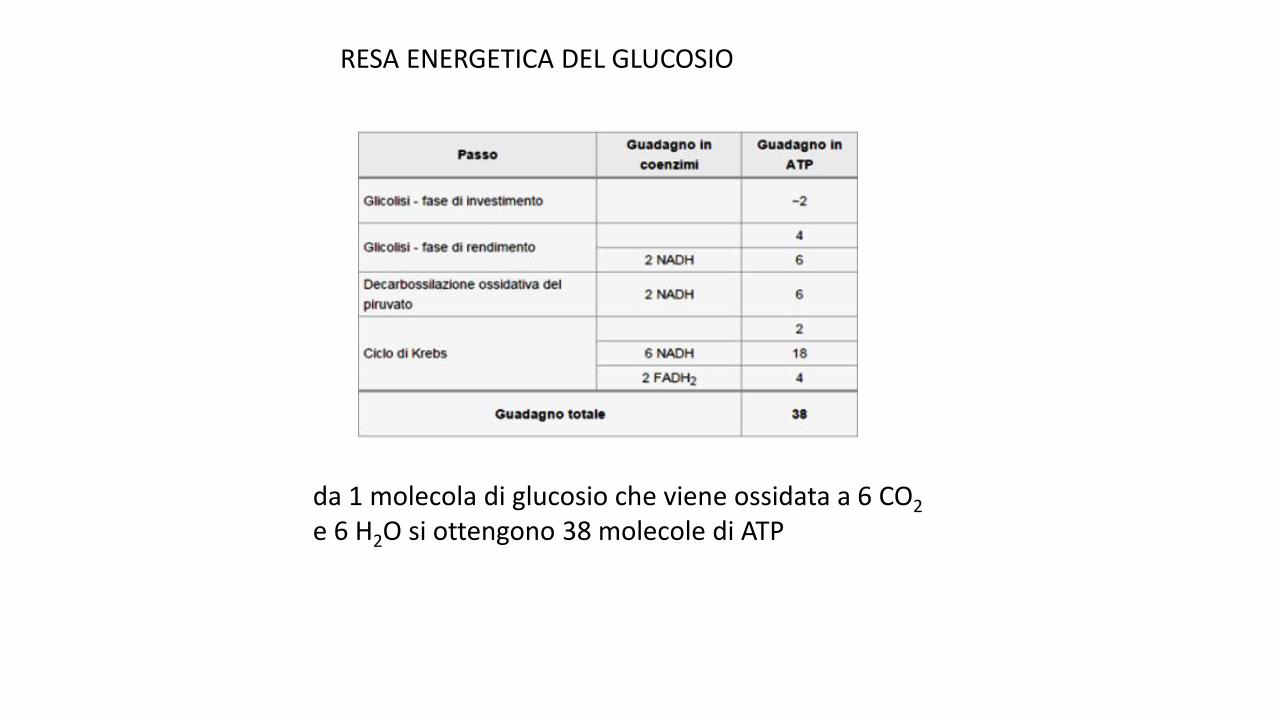
da 1 molecola di glucosio che viene ossidata a 6 CO2
e 6 H2O si ottengono 38 molecole di ATP
RESA ENERGETICA DEL GLUCOSIO

il piruvato prodotto nel citosol al termine della glicolisi può avere vari destini
trasporto nel mitocondrio
+ CO2
decarbossilazione ossidativa(irreversibile)
~SCoA
NADHCoASH
al ciclo di Krebs
in condizioni anaerobichenel citoplasma
OH
C
H
C
O–
OCH3
lattato
il piruvato si riduce alattato perché funge daaccettore di elettroni,riossidando il NADH aNAD+ al posto di O2
viene poi riossidato a NAD+ a spese di O2
con produzione di ATP
alanina
alla sintesi di amminoacidi
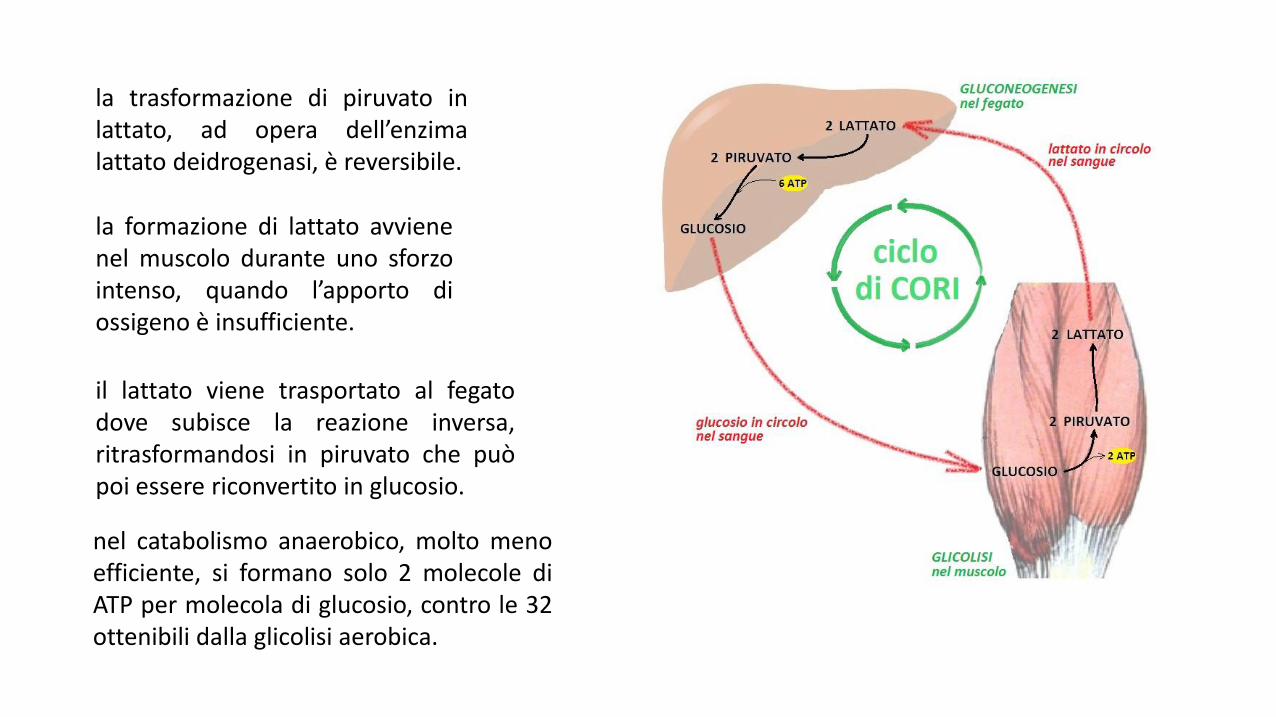
la trasformazione di piruvato inlattato, ad opera dell’enzimalattato deidrogenasi, è reversibile.
la formazione di lattato avvienenel muscolo durante uno sforzointenso, quando l’apporto diossigeno è insufficiente.
il lattato viene trasportato al fegatodove subisce la reazione inversa,ritrasformandosi in piruvato che puòpoi essere riconvertito in glucosio.
nel catabolismo anaerobico, molto menoefficiente, si formano solo 2 molecole diATP per molecola di glucosio, contro le 32ottenibili dalla glicolisi aerobica.
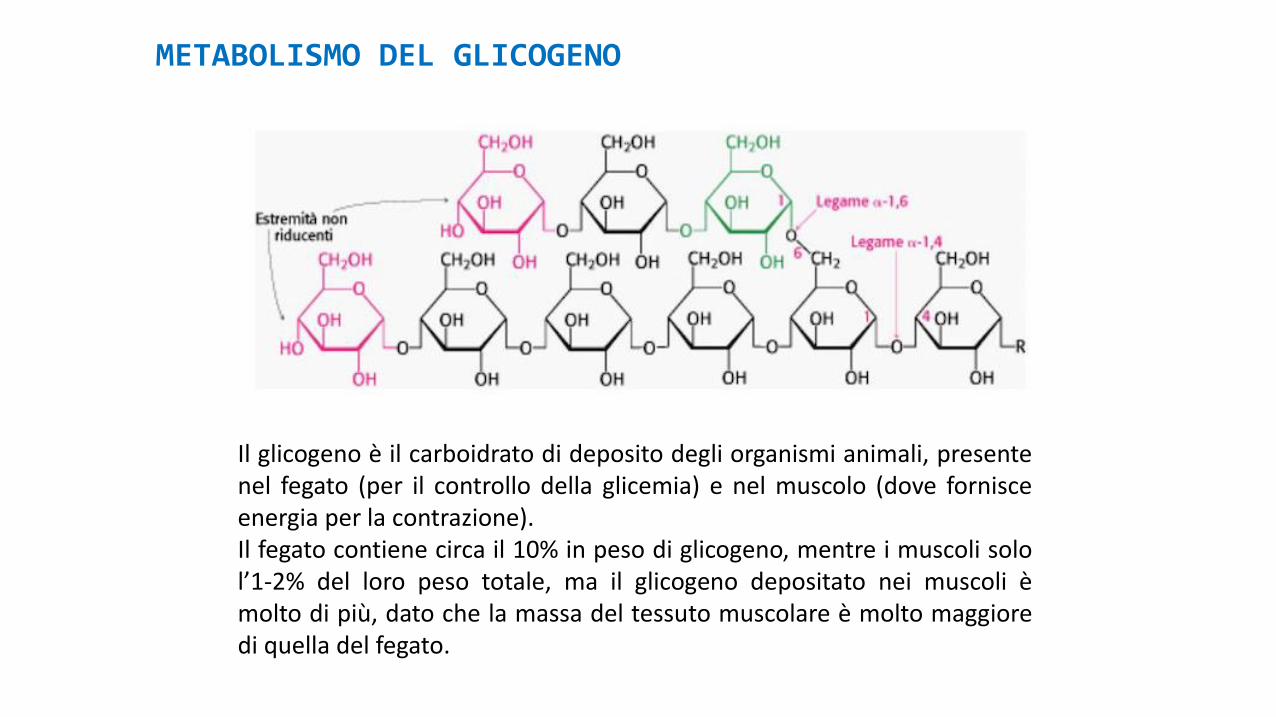
METABOLISMO DEL GLICOGENO
Il glicogeno è il carboidrato di deposito degli organismi animali, presentenel fegato (per il controllo della glicemia) e nel muscolo (dove fornisceenergia per la contrazione).Il fegato contiene circa il 10% in peso di glicogeno, mentre i muscoli solol’1-2% del loro peso totale, ma il glicogeno depositato nei muscoli èmolto di più, dato che la massa del tessuto muscolare è molto maggioredi quella del fegato.
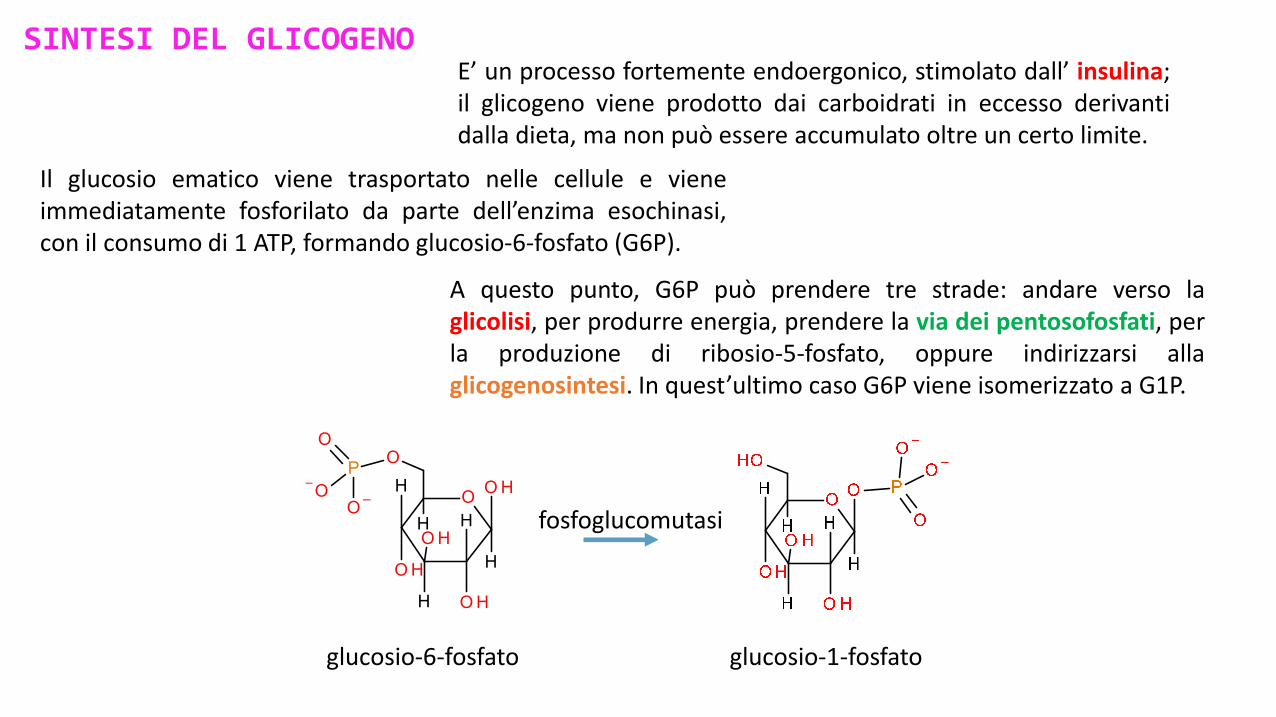
SINTESI DEL GLICOGENOE’ un processo fortemente endoergonico, stimolato dall’ insulina;il glicogeno viene prodotto dai carboidrati in eccesso derivantidalla dieta, ma non può essere accumulato oltre un certo limite.
P
O–O
–
OO
OH
OH
OH
H
H
H
H
O
H
OH
glucosio-6-fosfato glucosio-1-fosfato
fosfoglucomutasi
Il glucosio ematico viene trasportato nelle cellule e vieneimmediatamente fosforilato da parte dell’enzima esochinasi,con il consumo di 1 ATP, formando glucosio-6-fosfato (G6P).
A questo punto, G6P può prendere tre strade: andare verso laglicolisi, per produrre energia, prendere la via dei pentosofosfati, perla produzione di ribosio-5-fosfato, oppure indirizzarsi allaglicogenosintesi. In quest’ultimo caso G6P viene isomerizzato a G1P.

G1P
uridina trifosfato(UTP)
uridin difosfoglucosio(UDPG)
+ PPi
glucosio—fosfato uridil transferasi
SINTESI DEL GLICOGENO
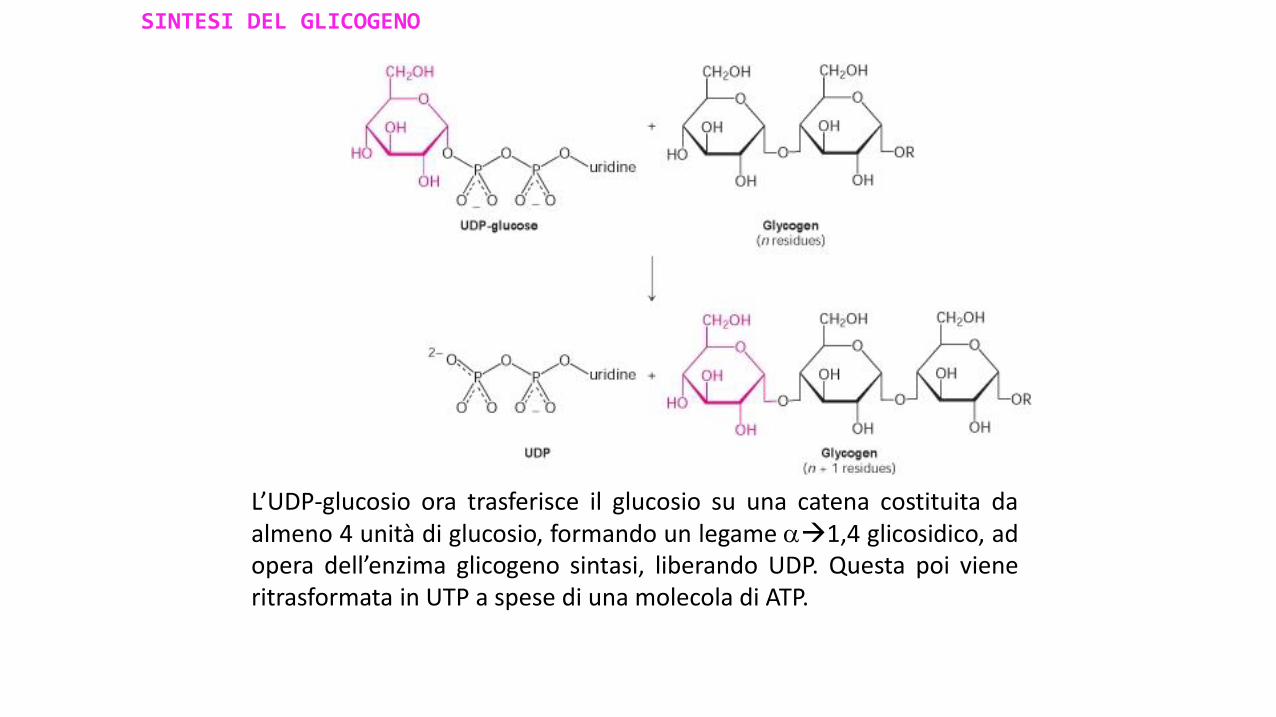
L’UDP-glucosio ora trasferisce il glucosio su una catena costituita daalmeno 4 unità di glucosio, formando un legame 1,4 glicosidico, adopera dell’enzima glicogeno sintasi, liberando UDP. Questa poi vieneritrasformata in UTP a spese di una molecola di ATP.
SINTESI DEL GLICOGENO

22
L’enzima ramificante poi staccadalla catena un frammento dialmeno 6 unità per legarlo, sullastessa catena o su un’altra, conun legame 1,6 glicosidico, anon meno di 4 unità di distanzada un’altra ramificazione.
Le ramificazioni rendono il glicogeno più solubile, e soprattuttodotato di un maggior numero di residui terminali, sui qualipossono agire gli enzimi glicogeno sintasi e glicogeno fosforilasi,aumentando la velocità sia di sintesi che di degradazione.
SINTESI DEL GLICOGENO

23
Il glicogeno è immagazzinato nel citosol. Grazie alla suastruttura molto ramificata è in grado di mobilizzare piùunità di glucosio idrolizzandosi rapidamente al bisogno, inseguito al segnale degli ormoni glucagone e adrenalina, adopera della glicogeno fosforilasi e dell’enzimaderamificante. In genere non viene demolito del tutto masolo ridotto di dimensioni.
GLICOGENOLISI

24
La glicogeno fosforilasi attacca l’unità di glucosio terminalestaccandole dalla catena sotto forma di glucosio-1-fosfato.
GLICOGENOLISI

25
La glicogeno fosforilasi stacca le unità di glucosio partendodall’estremità di tutte le ramificazioni. Quando arriva a 4 residui dallaramificazione, l’enzima deramificante trasferisce un frammento di 3unità di glucosio all’estremità di un’altra catena, e infine idrolizzal’ultimo legame 16 glicosidico, rilasciando glucosio non fosforilato.
GLICOGENOLISI
26
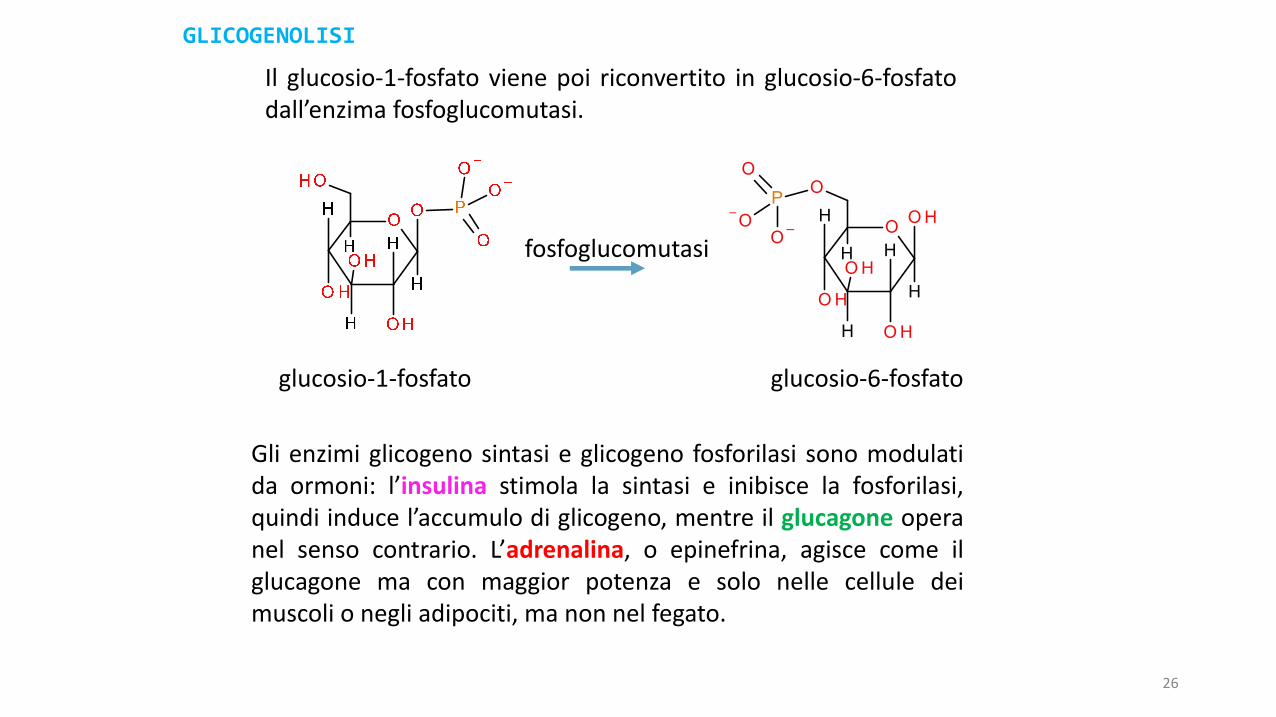
Gli enzimi glicogeno sintasi e glicogeno fosforilasi sono modulatida ormoni: l’insulina stimola la sintasi e inibisce la fosforilasi,quindi induce l’accumulo di glicogeno, mentre il glucagone operanel senso contrario. L’adrenalina, o epinefrina, agisce come ilglucagone ma con maggior potenza e solo nelle cellule deimuscoli o negli adipociti, ma non nel fegato.
Il glucosio-1-fosfato viene poi riconvertito in glucosio-6-fosfatodall’enzima fosfoglucomutasi.
P
O–O
–
OO
OH
OH
OH
H
H
H
H
O
H
OH
glucosio-6-fosfatoglucosio-1-fosfato
fosfoglucomutasi
GLICOGENOLISI

27
P
O–O
–
OO
OH
OH
OH
H
H
H
H
O
H
OH
glucosio-6-fosfatonel muscolo
GLICOLISIproduzione dipiruvato e ATP
nel fegato
OHO
OH
H H
OH
OH
H
H
OH H
la glucosio-6-fosfatasi stacca Pie libera glucosio che in questaforma può uscire dalle cellule epassare nel sangue
questo è il meccanismomediante il quale il fegatoregola la glicemia
28
VIA DEI PENTOSOFOSFATI
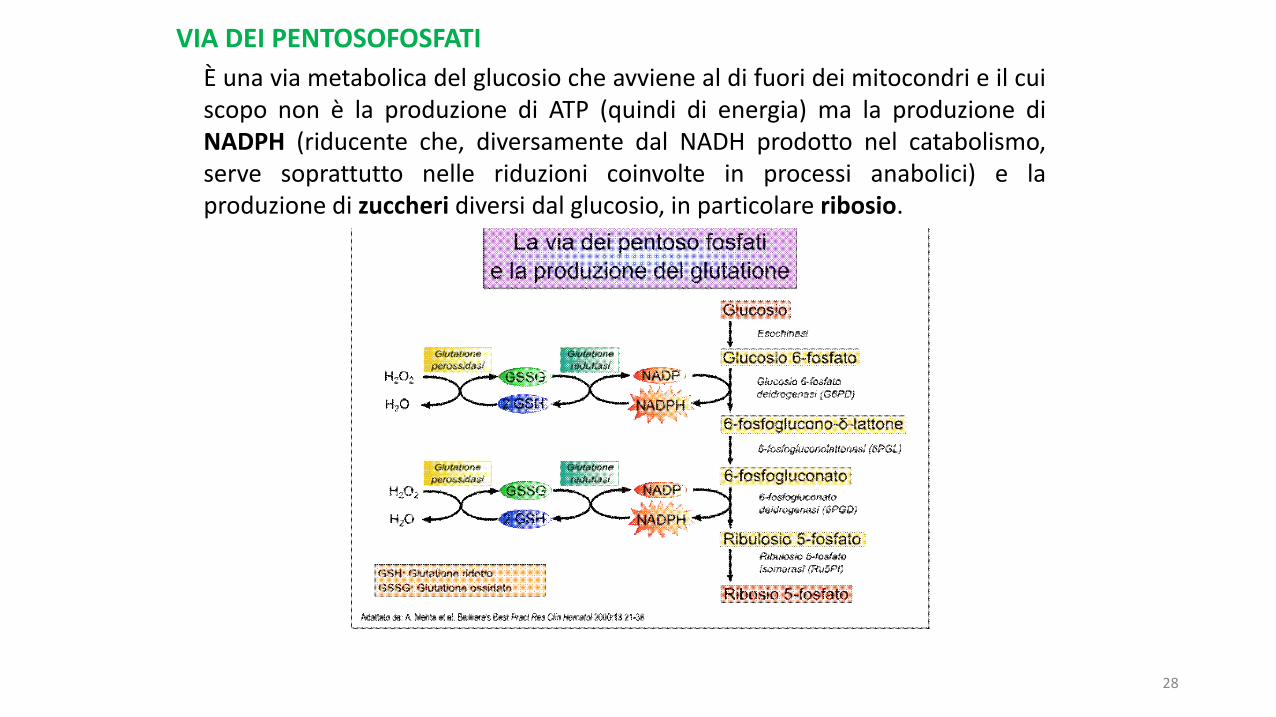
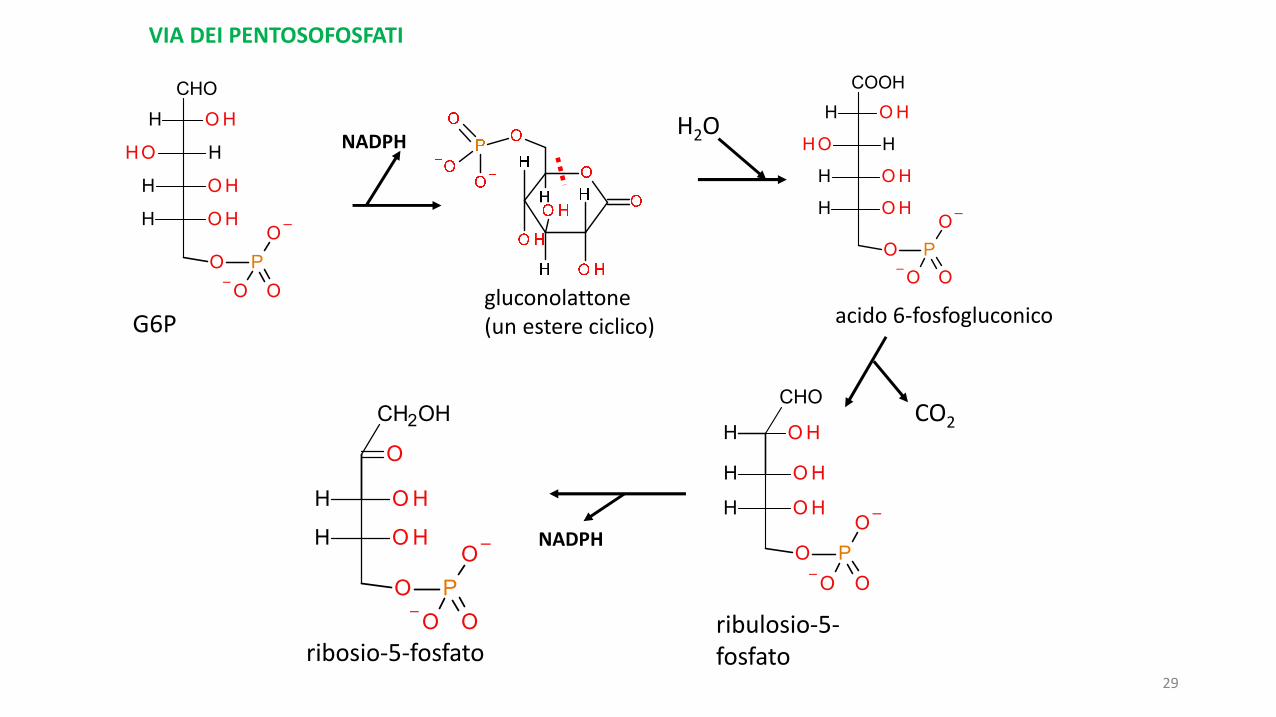
È una via metabolica del glucosio che avviene al di fuori dei mitocondri e il cuiscopo non è la produzione di ATP (quindi di energia) ma la produzione diNADPH (riducente che, diversamente dal NADH prodotto nel catabolismo,serve soprattutto nelle riduzioni coinvolte in processi anabolici) e laproduzione di zuccheri diversi dal glucosio, in particolare ribosio.
29
NADPH
COOH
P
O–
O
O–
O
OHH
H
H
OH
OH
OHHH2O
gluconolattone(un estere ciclico) acido 6-fosfogluconico
ribulosio-5-fosfato
NADPH
CH2OH
H OH
P
O–
O
O–
O
OHH
O
CO2
CHO
OHH
HOH
H OH
P
O–
O
O–
O
OHH
G6P
CHO
H
H OH
P
O–
O
O–
O
OHH
OH
ribosio-5-fosfato
VIA DEI PENTOSOFOSFATI
30
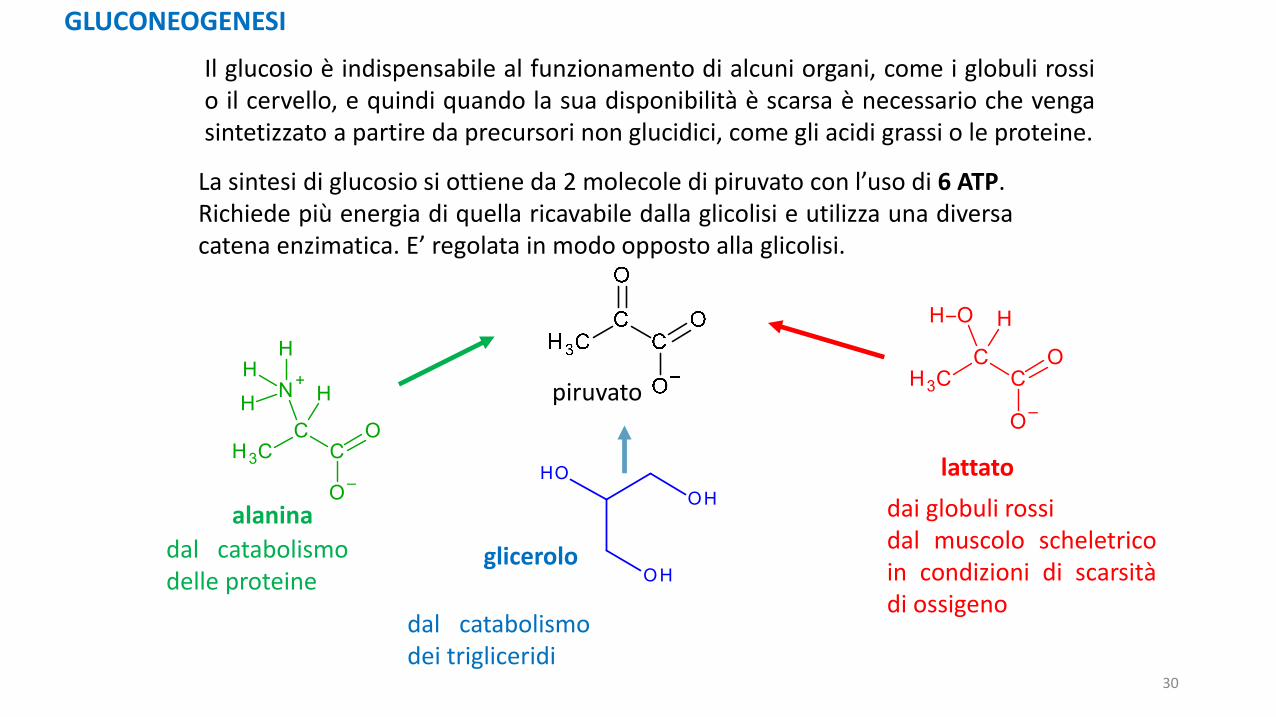
GLUCONEOGENESI
Il glucosio è indispensabile al funzionamento di alcuni organi, come i globuli rossio il cervello, e quindi quando la sua disponibilità è scarsa è necessario che vengasintetizzato a partire da precursori non glucidici, come gli acidi grassi o le proteine.
La sintesi di glucosio si ottiene da 2 molecole di piruvato con l’uso di 6 ATP.Richiede più energia di quella ricavabile dalla glicolisi e utilizza una diversacatena enzimatica. E’ regolata in modo opposto alla glicolisi.
piruvato
OH
OH
OH
glicerolo
C
HN+
HH
H
C
O–
OCH3
alanina
dal catabolismodelle proteine
dal catabolismodei trigliceridi
C
HOH
C
O–
OCH3
lattato
dai globuli rossidal muscolo scheletricoin condizioni di scarsitàdi ossigeno

glucosio
piruvato
alanina
2 ATP
NH2
MUSCOLO
glucosio
lattato
urea
2 ATP
GLOBULI ROSSI
glicolisi
glicolisi
glucosio
piruvato
alanina
6 ATP
NH2
FEGATOgl
uco
neo
gen
esi
lattato
ciclo dell’urea 4 ATP
i prodotti non completamente ossidati del catabolismo,presenti nei muscoli e nei globuli rossi, tornano al fegato peressere riciclati, ma il bilancio energetico è negativo.
32
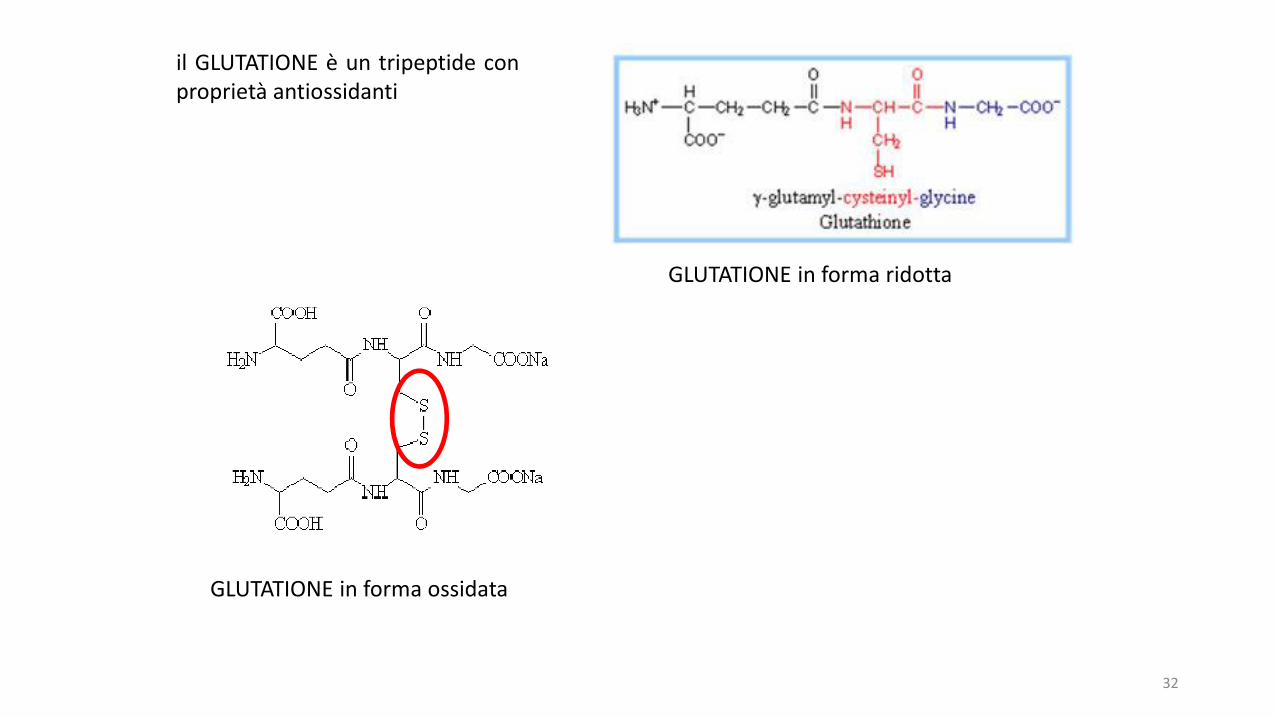
il GLUTATIONE è un tripeptide conproprietà antiossidanti
GLUTATIONE in forma ridotta
GLUTATIONE in forma ossidata

33
METABOLISMO DELL’ETANOLO
NADH NADH
Questa via metabolica procede mediante gli enzimi alcool deidrogenasi ealdeide deidrogenasi; il NADH che ne deriva è poi convertito in ATP, per unaresa energetica di circa 7 kcal/mol di alcool. E’ una via metabolica veloce, chepuò fornire molte calorie al giorno, ma l’acetaldeide è tossica perché è ingrado di formare complessi con proteine e acidi nucleici. Inoltre l’eccessivaproduzione di NADH stimola la produzione di lattato, con conseguenteacidosi, e inibisce la gluconeogenesi, con conseguente ipoglicemia.

34
IN CONCLUSIONE:
VIA DEI PENTOSOFOSFATI:• sintesi di ribosio-5-fosfato• NADPH + H+ da usare in processi anabolici• sintesi di altri zuccheri
GLICOLISI:• aerobica – da 1 mol di glucosio 2 mol piruvato + 2 ATP + 2 NADH + H+
• anaerobica - il piruvato viene ridotto a lattato a spese di NADH (ciclo di Cori)
GLICOGENO:• glicogenosintesi• glicogenolisi
GLUCONEOGENESI:produzione di piruvato da avviare alla glicolisi a partire da
• alanina (dal catabolismo delle proteine)• lattato (dalla glicolisi anaerobica)• glicerolo (dal catabolismo dei trigliceridi)
G6P







