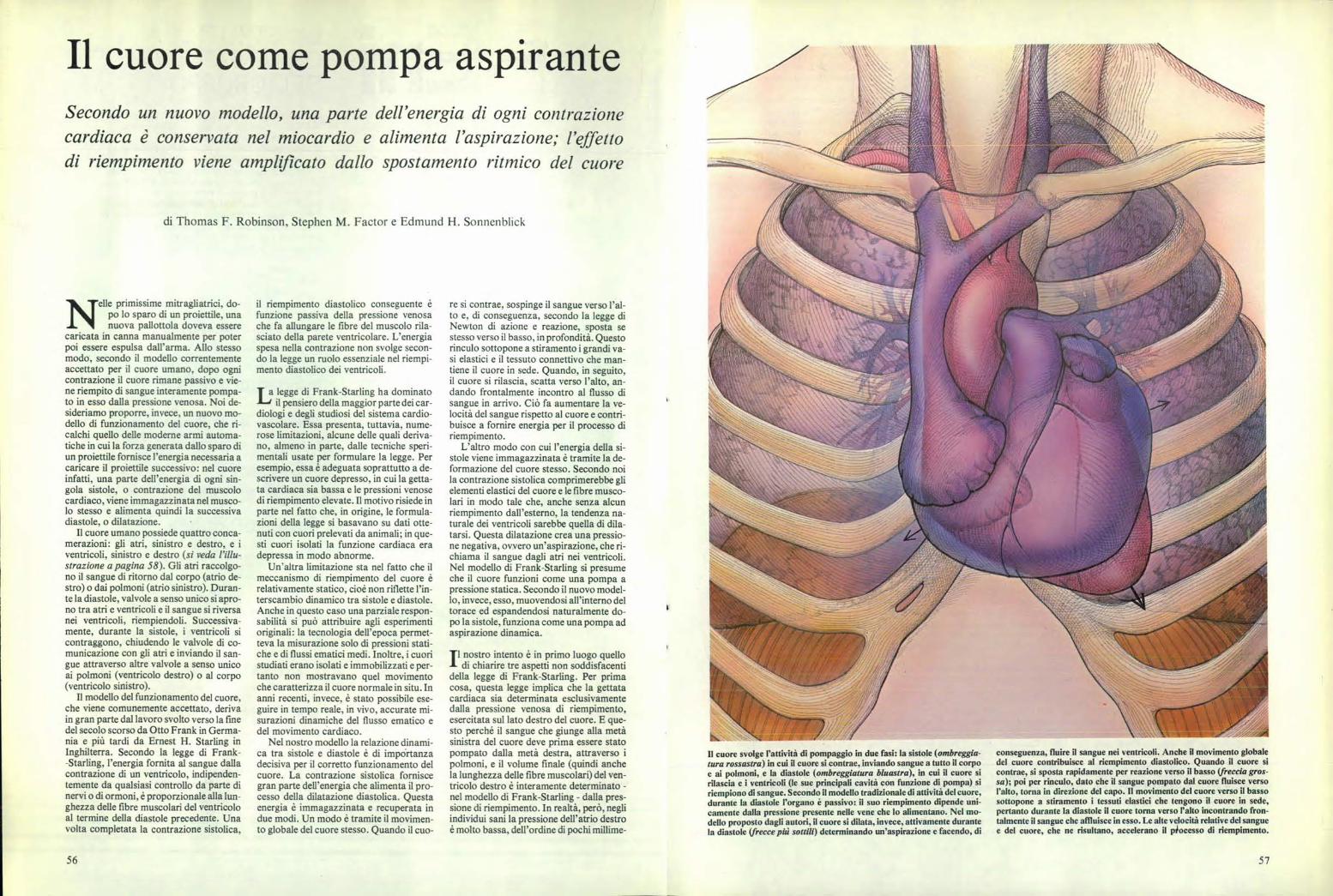
Il cuore svolge l'attività di pompaggio in due fasi: la sistole (ombreggia- tura rossastra) in cui il cuore si contrae, inviando sangue a tutto il corpo e ai polmoni, e la diastole (ombreggiatura bluastra), in cui il cuore si rilascia e i ventricoli (le sue principali cavità con funzione di pompa) si riempiono di sangue. Secondo il modello tradizionale di attività del cuore, durante la diastole l'organo è passivo: il suo riempimento dipende uni- camente dalla pressione presente nelle vene che lo alimentano. Nel mo- dello proposto dagli autori, il cuore si dilata, invece, attivamente durante la diastole (frecce più sottili) determinando un'aspirazione e facendo, di conseguenza, fluire il sangue nei ventricoli. Anche il movimento globale del cuore contribuisce al riempimento diastolico. Quando il cuore si contrae, si sposta rapidamente per reazione verso il basso (freccia gros- sa); poi per rinculo, dato che il sangue pompato dal cuore fluisce verso l'alto, torna in direzione del capo. Il movimento del cuore verso il basso sottopone a stiramento i tessuti elastici che tengono il cuore in sede, pertanto durante la diastole il cuore torna verso l'alto incontrando fron- talmente il sangue che affluisce in esso. Le alte velocità relative del sangue e del cuore, che ne risultano, accelerano il piocesso di riempimento. Il cuore come pompa aspirante Secondo un nuovo modello, una parte dell'energia di ogni contrazione cardiaca è conservata nel miocardio e alimenta l'aspirazione; l'effetto di riempimento viene amplificato dallo spostamento ritmico del cuore di Thomas F. Robinson. Stephen M. Factor e Edmund H. Sonnenblick s elle primissime mitragliatrici, do- po lo sparo di un proiettile, una nuova pallottola doveva essere caricata in canna manualmente per poter poi essere espulsa dall'arma. Allo stesso modo, secondo il modello correntemente accettato per il cuore umano, dopo ogni contrazione il cuore rimane passivo e vie- ne riempito di sangue interamente pompa- to in esso dalla pressione venosa. Noi de- sideriamo proporre, invece, un nuovo mo- dello di funzionamento del cuore, che ri- calchi quello delle moderne armi automa- tiche in cui la forza generata dallo sparo di un proiettile fornisce l'energia necessaria a caricare il proiettile successivo: nel cuore infatti, una parte dell'energia di ogni sin- gola sistole, o contrazione del muscolo cardiaco, viene immagazzinata nel musco- lo stesso e alimenta quindi la successiva diastole, o dilatazione. Il cuore umano possiede quattro conca- merazioni: gli atri, sinistro e destro, e i ventricoli, sinistro e destro (si veda l'illu- strazione a pagina 58). Gli atri raccolgo- no il sangue di ritorno dal corpo (atrio de- stro) o dai polmoni (atrio sinistro). Duran- te la diastole, valvole a senso unico si apro- no tra atri e ventricoli e il sangue si riversa nei ventricoli, riempiendoli. Successiva- mente, durante la sistole, i ventricoli si contraggono, chiudendo le valvole di co- municazione con gli atri e inviando il san- gue attraverso altre valvole a senso unico ai polmoni (ventricolo destro) o al corpo (ventricolo sinistro). Il modello del funzionamento del cuore, che viene comunemente accettato, deriva in gran parte dal lavoro svolto verso la fine del secolo scorso da Otto Frank in Germa- nia e più tardi da Ernest H. Starling in Inghilterra. Secondo la legge di Frank- -Starling, l'energia fornita al sangue dalla contrazione di un ventricolo, indipenden- temente da qualsiasi controllo da parte di nervi o di ormoni, è proporzionale alla lun- ghezza delle fibre muscolari del ventricolo al termine della diastole precedente. Una volta completata la contrazione sistolica, il riempimento diastolico conseguente è funzione passiva della pressione venosa che fa allungare le fibre del muscolo rila- sciato della parete ventricolare. L'energia spesa nella contrazione non svolge secon- do la legge un ruolo essenziale nel riempi- mento diastolico dei ventricoli. T a legge di Frank-Starling ha dominato il pensiero della maggior parte dei car- diologi e degli studiosi del sistema cardio- vascolare. Essa presenta, tuttavia, nume- rose limitazioni, alcune delle quali deriva- no, almeno in parte, dalle tecniche speri- mentali usate per formulare la legge. Per esempio, essa è adeguata soprattutto a de- scrivere un cuore depresso, in cui la getta- ta cardiaca sia bassa e le pressioni venose di riempimento elevate. Il motivo risiede in parte nel fatto che, in origine, le formula- zioni della legge si basavano su dati otte- nuti con cuori prelevati da animali; in que- sti cuori isolati la funzione cardiaca era depressa in modo abnorme. Un'altra limitazione sta nel fatto che il meccanismo di riempimento del cuore è relativamente statico, cioè non riflette l'in- terscambio dinamico tra si stole e diastole. Anche in questo caso una parziale respon- sabilità si può attribuire agli esperimenti originali: la tecnologia dell'epoca permet- teva la misurazione solo di pressioni stati- che e di flussi ematici medi. Inoltre, i cuori studiati erano isolati e immobilizzati e per- tanto non mostravano quel movimento che caratterizza il cuore normale in situ. In anni recenti, invece, è stato possibile ese- guire in tempo reale, in vivo, accurate mi- surazioni dinamiche del flusso ematico e del movimento cardiaco. Nel nostro modello la relazione dinami- ca tra sistole e diastole è di importanza decisiva per il corretto funzionamento del cuore. La contrazione sistolica fornisce gran parte dell'energia che alimenta il pro- cesso della dilatazione diastolica. Questa energia è immagazzinata e recuperata in due modi. Un modo è tramite il movimen- to globale del cuore stesso. Quando il cuo- re si contrae, sospinge il sangue verso l'al- to e, di conseguenza, secondo la legge di Newton di azione e reazione, sposta se stesso verso il basso, in profondità. Questo rinculo sottopone a stiramento i grandi va- si elastici e il tessuto connettivo che man- tiene il cuore in sede. Quando, in seguito, il cuore si rilascia, scatta verso l'alto, an- dando frontalmente incontro al flusso di sangue in arrivo. Ciò fa aumentare la ve- locità del sangue rispetto al cuore e contri- buisce a fornire energia per il processo di riempimento. L'altro modo con cui l'energia della si- stole viene immagazzinata è tramite la de- formazione del cuore stesso. Secondo noi la contrazione sistolica comprimerebbe gli elementi elastici del cuore e le fibre musco- lari in modo tale che, anche senza alcun riempimento dall'esterno, la tendenza na- turale dei ventricoli sarebbe quella di dila- tarsi. Questa dilatazione crea una pressio- ne negativa, ovvero un'aspirazione, che ri- chiama il sangue dagli atri nei ventricoli. Nel modello di Frank-Starling si presume che il cuore funzioni come una pompa a pressione statica. Secondo il nuovo model- lo, invece, esso, muovendosi all'interno del torace ed espandendosi naturalmente do- po la sistole, funziona come una pompa ad aspirazione dinamica. Tl nostro intento è in primo luogo quello di chiarire tre aspetti non soddisfacenti della legge di Frank-Starling. Per prima cosa, questa legge implica che la gettata cardiaca sia determinata esclusivamente dalla pressione venosa di riempimento, esercitata sul lato destro del cuore. E que- sto perché il sangue che giunge alla metà sinistra del cuore deve prima essere stato pompato dalla metà destra, attraverso i polmoni, e il volume finale (quindi anche la lunghezza delle fibre muscolari) del ven- tricolo destro è interamente determinato - nel modello di Frank-Starling - dalla pres- sione di riempimento. In realtà, però, negli individui sani la pressione dell'atrio destro è molto bassa, dell'ordine di pochi minime- 56 57
Transcript
Il cuore svolge l'attività di pompaggio in due fasi: la sistole (ombreggia-tura rossastra) in cui il cuore si contrae, inviando sangue a tutto il corpoe ai polmoni, e la diastole (ombreggiatura bluastra), in cui il cuore sirilascia e i ventricoli (le sue principali cavità con funzione di pompa) siriempiono di sangue. Secondo il modello tradizionale di attività del cuore,durante la diastole l'organo è passivo: il suo riempimento dipende uni-camente dalla pressione presente nelle vene che lo alimentano. Nel mo-dello proposto dagli autori, il cuore si dilata, invece, attivamente durantela diastole (frecce più sottili) determinando un'aspirazione e facendo, di
conseguenza, fluire il sangue nei ventricoli. Anche il movimento globaledel cuore contribuisce al riempimento diastolico. Quando il cuore sicontrae, si sposta rapidamente per reazione verso il basso (freccia gros-sa); poi per rinculo, dato che il sangue pompato dal cuore fluisce versol'alto, torna in direzione del capo. Il movimento del cuore verso il bassosottopone a stiramento i tessuti elastici che tengono il cuore in sede,pertanto durante la diastole il cuore torna verso l'alto incontrando fron-talmente il sangue che affluisce in esso. Le alte velocità relative del sanguee del cuore, che ne risultano, accelerano il piocesso di riempimento.
Il cuore come pompa aspiranteSecondo un nuovo modello, una parte dell'energia di ogni contrazionecardiaca è conservata nel miocardio e alimenta l'aspirazione; l'effettodi riempimento viene amplificato dallo spostamento ritmico del cuore
di Thomas F. Robinson. Stephen M. Factor e Edmund H. Sonnenblick
s
elle primissime mitragliatrici, do-po lo sparo di un proiettile, unanuova pallottola doveva essere
caricata in canna manualmente per poterpoi essere espulsa dall'arma. Allo stessomodo, secondo il modello correntementeaccettato per il cuore umano, dopo ognicontrazione il cuore rimane passivo e vie-ne riempito di sangue interamente pompa-to in esso dalla pressione venosa. Noi de-sideriamo proporre, invece, un nuovo mo-dello di funzionamento del cuore, che ri-calchi quello delle moderne armi automa-tiche in cui la forza generata dallo sparo diun proiettile fornisce l'energia necessaria acaricare il proiettile successivo: nel cuoreinfatti, una parte dell'energia di ogni sin-gola sistole, o contrazione del muscolocardiaco, viene immagazzinata nel musco-lo stesso e alimenta quindi la successivadiastole, o dilatazione.
Il cuore umano possiede quattro conca-merazioni: gli atri, sinistro e destro, e iventricoli, sinistro e destro (si veda l'illu-strazione a pagina 58). Gli atri raccolgo-no il sangue di ritorno dal corpo (atrio de-stro) o dai polmoni (atrio sinistro). Duran-te la diastole, valvole a senso unico si apro-no tra atri e ventricoli e il sangue si riversanei ventricoli, riempiendoli. Successiva-mente, durante la sistole, i ventricoli sicontraggono, chiudendo le valvole di co-municazione con gli atri e inviando il san-gue attraverso altre valvole a senso unicoai polmoni (ventricolo destro) o al corpo(ventricolo sinistro).
Il modello del funzionamento del cuore,che viene comunemente accettato, derivain gran parte dal lavoro svolto verso la finedel secolo scorso da Otto Frank in Germa-nia e più tardi da Ernest H. Starling inInghilterra. Secondo la legge di Frank--Starling, l'energia fornita al sangue dallacontrazione di un ventricolo, indipenden-temente da qualsiasi controllo da parte dinervi o di ormoni, è proporzionale alla lun-ghezza delle fibre muscolari del ventricoloal termine della diastole precedente. Unavolta completata la contrazione sistolica,
il riempimento diastolico conseguente èfunzione passiva della pressione venosache fa allungare le fibre del muscolo rila-sciato della parete ventricolare. L'energiaspesa nella contrazione non svolge secon-do la legge un ruolo essenziale nel riempi-mento diastolico dei ventricoli.
T a legge di Frank-Starling ha dominatoil pensiero della maggior parte dei car-
diologi e degli studiosi del sistema cardio-vascolare. Essa presenta, tuttavia, nume-rose limitazioni, alcune delle quali deriva-no, almeno in parte, dalle tecniche speri-mentali usate per formulare la legge. Peresempio, essa è adeguata soprattutto a de-scrivere un cuore depresso, in cui la getta-ta cardiaca sia bassa e le pressioni venosedi riempimento elevate. Il motivo risiede inparte nel fatto che, in origine, le formula-zioni della legge si basavano su dati otte-nuti con cuori prelevati da animali; in que-sti cuori isolati la funzione cardiaca eradepressa in modo abnorme.
Un'altra limitazione sta nel fatto che ilmeccanismo di riempimento del cuore èrelativamente statico, cioè non riflette l'in-terscambio dinamico tra si stole e diastole.Anche in questo caso una parziale respon-sabilità si può attribuire agli esperimentioriginali: la tecnologia dell'epoca permet-teva la misurazione solo di pressioni stati-che e di flussi ematici medi. Inoltre, i cuoristudiati erano isolati e immobilizzati e per-tanto non mostravano quel movimentoche caratterizza il cuore normale in situ. Inanni recenti, invece, è stato possibile ese-guire in tempo reale, in vivo, accurate mi-surazioni dinamiche del flusso ematico edel movimento cardiaco.
Nel nostro modello la relazione dinami-ca tra sistole e diastole è di importanzadecisiva per il corretto funzionamento delcuore. La contrazione sistolica forniscegran parte dell'energia che alimenta il pro-cesso della dilatazione diastolica. Questaenergia è immagazzinata e recuperata indue modi. Un modo è tramite il movimen-to globale del cuore stesso. Quando il cuo-
re si contrae, sospinge il sangue verso l'al-to e, di conseguenza, secondo la legge diNewton di azione e reazione, sposta sestesso verso il basso, in profondità. Questorinculo sottopone a stiramento i grandi va-si elastici e il tessuto connettivo che man-tiene il cuore in sede. Quando, in seguito,il cuore si rilascia, scatta verso l'alto, an-dando frontalmente incontro al flusso disangue in arrivo. Ciò fa aumentare la ve-locità del sangue rispetto al cuore e contri-buisce a fornire energia per il processo diriempimento.
L'altro modo con cui l'energia della si-stole viene immagazzinata è tramite la de-formazione del cuore stesso. Secondo noila contrazione sistolica comprimerebbe glielementi elastici del cuore e le fibre musco-lari in modo tale che, anche senza alcunriempimento dall'esterno, la tendenza na-turale dei ventricoli sarebbe quella di dila-tarsi. Questa dilatazione crea una pressio-ne negativa, ovvero un'aspirazione, che ri-chiama il sangue dagli atri nei ventricoli.Nel modello di Frank-Starling si presumeche il cuore funzioni come una pompa apressione statica. Secondo il nuovo model-lo, invece, esso, muovendosi all'interno deltorace ed espandendosi naturalmente do-po la sistole, funziona come una pompa adaspirazione dinamica.
Tl nostro intento è in primo luogo quellodi chiarire tre aspetti non soddisfacenti
della legge di Frank-Starling. Per primacosa, questa legge implica che la gettatacardiaca sia determinata esclusivamentedalla pressione venosa di riempimento,esercitata sul lato destro del cuore. E que-sto perché il sangue che giunge alla metàsinistra del cuore deve prima essere statopompato dalla metà destra, attraverso ipolmoni, e il volume finale (quindi anchela lunghezza delle fibre muscolari) del ven-tricolo destro è interamente determinato -nel modello di Frank-Starling - dalla pres-sione di riempimento. In realtà, però, negliindividui sani la pressione dell'atrio destroè molto bassa, dell'ordine di pochi minime-
56
57
VALVOLEA SENSO
UNICO
ATRIOSINISTRO
VALVOLAA SENSOUNICO
VENTRICOLOSINISTROVENTRICOLO
DESTRO
ARTERIAPOLMONARE
ATRIODESTRO
VALVOLAA SENSO
UNICO
Il cuore umano possiede quattro cavità. L'atrio destro raccoglie il sangue privo di ossigeno pro-veniente dal resto del corpo e lo invia, attraverso una valvola a senso unico, nel ventricolo destro.Questo, attraverso un'altra valvola, lo pompa verso i polmoni per mezzo dell'arteria polmonare.Il sangue ossigenato che ritorna dai polmoni si raccoglie nell'atrio sinistro prima di riempire ilventricolo sinistro. Quest'ultimo poi pompa il sangue ossigenato nel corpo mediante l'aorta.
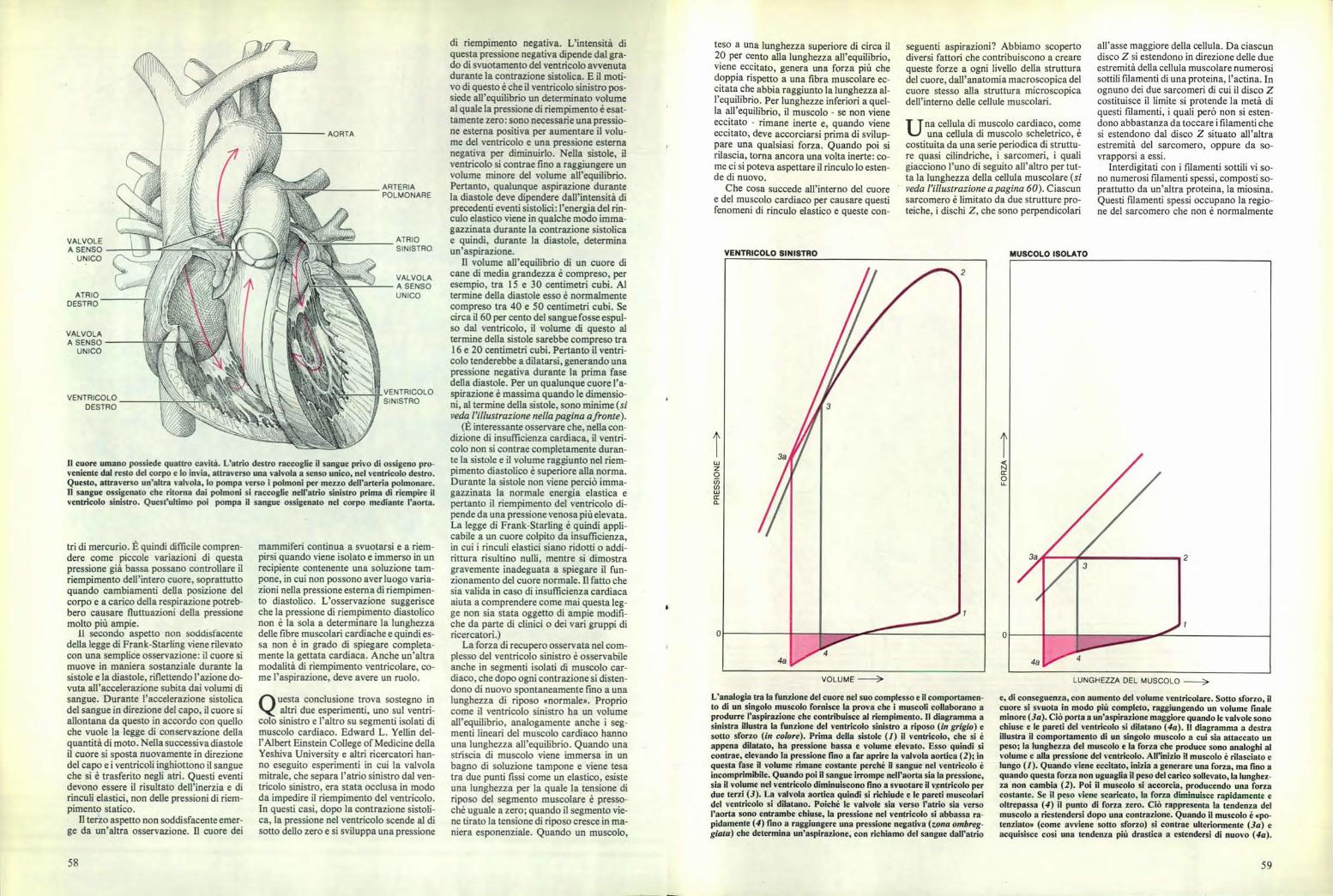
VENTRICOLO SINISTRO MUSCOLO ISOLATO
VOLUME
L'analogia tra la funzione del cuore nel suo complesso e il comportamen-to di un singolo muscolo fornisce la prova che i muscoli collaborano aprodurre l'aspirazione che contribuisce al riempimento. Il diagramma asinistra illustra la funzione del ventricolo sinistro a riposo (in grigio) esotto sforzo (in colore). Prima della sistole (1) il ventricolo, che si èappena dilatato, ha pressione bassa e volume elevato. Esso quindi sicontrae, elevando la pressione fino a far aprire la valvola aortica (2); inquesta fase il volume rimane costante perché il sangue nel ventricolo èincomprimibile. Quando poi il sangue irrompe nell'aorta sia la pressione,sia il volume nel ventricolo diminuiscono fino a svuotare il ventricolo perdue terzi (3). La valvola aortica quindi si richiude e le pareti muscolaridel ventricolo si dilatano. Poiché le valvole sia verso l'atrio sia versol'aorta sono entrambe chiuse, la pressione nel ventricolo si abbassa ra-pidamente (4) fino a raggiungere una pressione negativa (zona ombreg-giata) che determina un'aspirazione, con richiamo del sangue dall'atrio
LUNGHEZZA DEL MUSCOLO
e, di conseguenza, con aumento del volume ventricolare. Sotto sforzo, ilcuore si svuota in modo più completo, raggiungendo un volume finaleminore (3a). Ciò porta a un'aspirazione maggiore quando le valvole sonochiuse e le pareti del ventricolo si dilatano (4a). Il diagramma a destraillustra il comportamento di un singolo muscolo a cui sia attaccato unpeso; la lunghezza del muscolo e la forza che produce sono analoghi alvolume e alla pressione del ventricolo. All'inizio il muscolo è rilasciato elungo (I). Quando viene eccitato, inizia a generare una forza, ma fino aquando questa forza non uguaglia il peso del carico sollevato, la lunghez-za non cambia (2). Poi il muscolo si accorcia, producendo una forzacostante. Se il peso viene scaricato, la forza diminuisce rapidamente eoltrepassa (4) il punto di forza zero. Ciò rappresenta la tendenza delmuscolo a riestendersi dopo una contrazione. Quando il muscolo è «po-tenziato» (come avviene sotto sforzo) si contrae ulteriormente (3a) eacquisisce così una tendenza più drastica a estendersi di nuovo (4a).
4a
N
Ou_
a
4a 4
tri di mercurio. È quindi difficile compren-dere come piccole variazioni di questapressione già bassa possano controllare ilriempimento dell'intero cuore, soprattuttoquando cambiamenti della posizione delcorpo e a carico della respirazione potreb-bero causare fluttuazioni della pressionemolto più ampie.
11 secondo aspetto non soddisfacentedella legge di Frank-Starling viene rilevatocon una semplice osservazione: il cuore simuove in maniera sostanziale durante lasistole e la diastole, riflettendo l'azione do-vuta all'accelerazione subita dai volumi disangue. Durante l'accelerazione sistolicadel sangue in direzione del capo, il cuore siallontana da questo in accordo con quelloche vuole la legge di conservazione dellaquantità di moto. Nella successiva diastoleil cuore si sposta nuovamente in direzionedel capo e i ventricoli inghiottono il sangueche si è trasferito negli atri. Questi eventidevono essere il risultato dell'inerzia e dirinculi elastici, non delle pressioni di riem-pimento statico.
Il terzo aspetto non soddisfacente emer-ge da un'altra osservazione. Il cuore dei
mammiferi continua a svuotarsi e a riem-pirsi quando viene isolato e immerso in unrecipiente contenente una soluzione tam-pone, in cui non possono aver luogo varia-zioni nella pressione esterna di riempimen-to diastolico. L'osservazione suggerisceche la pressione di riempimento diastoliconon è la sola a determinare la lunghezzadelle fibre muscolari cardiache e quindi es-sa non è in grado di spiegare completa-mente la gettata cardiaca. Anche un'altramodalità di riempimento ventricolare, co-me l'aspirazione, deve avere un ruolo.
Questa conclusione trova sostegno inaltri due esperimenti, uno sul ventri-
colo sinistro e l'altro su segmenti isolati dimuscolo cardiaco. Edward L. Yellin del-l'Albert Einstein College of Medicine dellaYeshiva University e altri ricercatori han-no eseguito esperimenti in cui la valvolamitrale, che separa l'atrio sinistro dal ven-tricolo sinistro, era stata occlusa in mododa impedire il riempimento del ventricolo.In questi casi, dopo la contrazione sistoli-ca, la pressione nel ventricolo scende al disotto dello zero e si sviluppa una pressione
di riempimento negativa. L'intensità diquesta pressione negativa dipende dal gra-do di svuotamento del ventricolo avvenutadurante la contrazione sistolica. E il moti-vo di questo è che il ventricolo sinistro pos-siede all'equilibrio un determinato volumeal quale la pressione di riempimento è esat-tamente zero: sono necessarie una pressio-ne esterna positiva per aumentare il volu-me del ventricolo e una pressione esternanegativa per diminuirlo. Nella sistole, ilventricolo si contrae fino a raggiungere unvolume minore del volume all'equilibrio.Pertanto, qualunque aspirazione durantela diastole deve dipendere dall'intensità diprecedenti eventi sistolici: l'energia del rin-culo elastico viene in qualche modo imma-gazzinata durante la contrazione sistolicae quindi, durante la diastole, determinaun'aspirazione.
Il volume all'equilibrio di un cuore dicane di media grandezza è compreso, peresempio, tra 15 e 30 centimetri cubi. Altermine della diastole esso è normalmentecompreso tra 40 e 50 centimetri cubi. Secirca il 60 per cento del sangue fosse espul-so dal ventricolo, il volume di questo altermine della sistole sarebbe compreso tra16 e 20 centimetri cubi. Pertanto il ventri-colo tenderebbe a dilatarsi, generando unapressione negativa durante la prima fasedella diastole. Per un qualunque cuore l'a-spirazione è massima quando le dimensio-ni, al termine della sistole, sono minime (siveda l'illustrazione nella pagina a fronte).
(È interessante osservare che, nella con-dizione di insufficienza cardiaca, il ventri-colo non si contrae completamente duran-te la sistole e il volume raggiunto nel riem-pimento diastolico è superiore alla norma.Durante la sistole non viene perciò imma-gazzinata la normale energia elastica epertanto il riempimento del ventricolo di-pende da una pressione venosa più elevata.La legge di Frank-Starling è quindi appli-cabile a un cuore colpito da insufficienza,in cui i rinculi elastici siano ridotti o addi-rittura risultino nulli, mentre si dimostragravemente inadeguata a spiegare il fun-zionamento del cuore normale. Il fatto chesia valida in caso di insufficienza cardiacaaiuta a comprendere come mai questa leg-ge non sia stata oggetto di ampie modifi-che da parte di clinici o dei vari gruppi diricercatori.)
La forza di recupero osservata nel com-plesso del ventricolo sinistro è osservabileanche in segmenti isolati di muscolo car-diaco, che dopo ogni contrazione si disten-dono di nuovo spontaneamente fino a unalunghezza di riposo «normale». Propriocome il ventricolo sinistro ha un volumeall'equilibrio, analogamente anche i seg-menti lineari del muscolo cardiaco hannouna lunghezza all'equilibrio. Quando unastriscia di muscolo viene immersa in unbagno di soluzione tampone e viene tesatra due punti fissi come un elastico, esisteuna lunghezza per la quale la tensione diriposo del segmento muscolare è presso-ché uguale a zero; quando il segmento vie-ne tirato la tensione di riposo cresce in ma-niera esponenziale. Quando un muscolo,
teso a una lunghezza superiore di circa il20 per cento alla lunghezza all'equilibrio,viene eccitato, genera una forza più chedoppia rispetto a una fibra muscolare ec-citata che abbia raggiunto la lunghezza al-l'equilibrio. Per lunghezze inferiori a quel-la all'equilibrio, il muscolo - se non vieneeccitato - rimane inerte e, quando vieneeccitato, deve accorciarsi prima di svilup-pare una qualsiasi forza. Quando poi sirilascia, torna ancora una volta inerte: co-me ci si poteva aspettare il rinculo lo esten-de di nuovo.
Che cosa succede all'interno del cuoree del muscolo cardiaco per causare questifenomeni di rinculo elastico e queste con-
seguenti aspirazioni? Abbiamo scopertodiversi fattori che contribuiscono a crearequeste forze a ogni livello della strutturadel cuore, dall'anatomia macroscopica delcuore stesso alla struttura microscopicadell'interno delle cellule muscolari.
Una cellula di muscolo cardiaco, comeuna cellula di muscolo scheletrico, è
costituita da una serie periodica di struttu-re quasi cilindriche, i sarcomeri, i qualigiacciono l'uno di seguito all'altro per tut-ta la lunghezza della cellula muscolare (siveda l'illustrazione a pagina 60). Ciascunsarcomero è limitato da due strutture pro-teiche, i dischi Z, che sono perpendicolari
all'asse maggiore della cellula. Da ciascundisco Z si estendono in direzione delle dueestremità della cellula muscolare numerosisottili filamenti di una proteina, l'actina. Inognuno dei due sarcomeri di cui il disco Zcostituisce il limite si protende la metà diquesti filamenti, i quali però non si esten-dono abbastanza da toccare i filamenti chesi estendono dal disco Z situato all'altraestremità del sarcomero, oppure da so-vrapporsi a essi.
Interdigitati con i filamenti sottili vi so-no numerosi filamenti spessi, composti so-prattutto da un'altra proteina, la miosina.Questi filamenti spessi occupano la regio-ne del sarcomero che non è normalmente
58 59
FILAMENTO SPESSO FILAMENTO SOTTILE DISCO Z
Una cellula muscolare cardiaca (in alto) è costituita in gran parte da unaschiera di fibre, ciascuna delle quali è formata da numerose unità. isarcomeri, collocate l'una di seguito all'altra. Un sarcomero (in basso,a) è delimitato a entrambe le estremità dai dischi Z. Filamenti sottili diuna proteina, l'actina, si estendono da ciascun disco Z verso il centro delsarcomero. Essi si interdigitano con filamenti spessi di miosina. un'altraproteina; questi secondi filamenti non raggiungono i dischi Z. La contra-zione del muscolo (b) ha luogo quando i filamenti sottili scorrono lungoi filamenti spessi verso il centro del sarcomero. avvicinando tra loro idischi Z. Allorché il muscolo è completamente contratto (c) essi, dipar-
tendosi dalle estremità opposte del sarcomero, si sovrappongono l'unoall'altro all'interno del graticcio costituito dai filamenti spessi. Ciò faaumentare la circonferenza della cellula muscolare e può sottoporre astiramento i dischi Z e altri elementi laterali dello scheletro cellulare (chenon compaiono nell'illustrazione); gli elementi citoscheletrici così stiratipossono fornire una forza di recupero che fa allungare la cellula musco-lare contratta precedentemente, diminuendone la circonferenza. Le cel-lule muscolari cardiache sono avvolte da una trama incrociata di tessutoconnettivo; nel cuore umano, altri trefoli di fibre connettive, i montanti,che sono ritorti come corde, fissano le cellule muscolari l'una all'altra.
TRAMACONNETTIVALE
a SARCOM ERO SARCOMERO
9.4AMONTANTE
raggiunta dai filamenti sottili e ciascun fi-lamento spesso si sovrappone ai filamentisottili provenienti dai due dischi Z che de-limitano il sarcomero. Quando il muscoloviene stimolato, i filamenti sottili prove-nienti da entrambi i dischi Z vengono tirati
lungo i filamenti spessi verso il centro delsarcomero. Questo movimento avvicina idischi Z, accorciando il sarcomero. È pro-prio la contrazione pressoché simultaneadei numerosi sarcomeri situati l'uno di se-guito all'altro all'interno della cellula che
offre una spiegazione alla notevole capa-cità delle cellule muscolari di accorciarsirapidamente.
Durante il ciclo cardiaco i sarcomeridelle cellule muscolari del cuore si accor-ciano da circa 2,05 micrometri (milionesi-
mi di metro) a 1,85 rnicrometri per riallun-garsi poi di nuovo a 2,05 micrometri. Co-me si ottiene questo allungamento? Unacerta frazione dell'energia di contrazionedeve essere immagazzinata nella cellulamuscolare, ma dove? Dati riguardanti lafisiologia suggeriscono che la causa del-l'allungamento debba ricercarsi sia nellastruttura dei sarcomeri, sia nelle compo-nenti della cellula muscolare che formanoil citoscheletro, cioè quell'insieme di ele-menti fibrosi interconnessi che contribui-scono al mantenimento della forma dellacellula.
I filamenti sottili all'interno del sarco-mero sono lunghi ciascuno circa un micro-metro e pertanto, allorché i sarcomeri sicontraggono, quelli provenienti dalle op-poste estremità di un sarcomero (vale adire i filamenti sottili che si estendono dadischi Z diversi) si incontrano al centro delsarcomero allorché questo raggiunge unalunghezza di circa 2 micrometri. Quandoil sarcomero si contrae ulteriormente, i fi-lamenti sottili che si estendono da dischi Zdiversi cominciano a sovrapporsi all'inter-no del graticcio costituito dai filamentispessi. In una simile condizione di «doppiasovrapposizione», il graticcio di filamentisi fa stipato e il sarcomero si espande dilato: la sua forma cilindrica aumenta dicirconferenza. L'espansione in senso late-rale stira i dischi Z e altri elementi struttu-rali laterali della cellula, fornendo un mec-canismo mediante il quale una parte del-l'energia di contrazione è immagazzinataper essere riutilizzata come energia diespansione.
Non è solo la microstruttura delle cellu-le muscolari a immagazzinare l'ener-
gia della contrazione sistolica. Sono stateraccolte prove sempre più numerose sulfatto che un ruolo importante viene svoltoanche dal tessuto connettivo che circondale singole cellule muscolari e dal tessutoche le collega.
Per esempio, trefoli di fibre connettivalisono avvolti a elica attorno alla superficieesterna di ciascuna cellula muscolare cilin-drica. Le fibre sono disposte in senso siaorario, sia antiorario, in modo da formareuna trama incrociata non dissimile daquella di una rete per amache. Le fibresono costituite soprattutto di collageno, u-na sostanza poco estensibile che resiste al-lo stiramento, e contengono inoltre quan-tità variabili di elastina. L'elastina è unaproteina extracellulare che possiede pro-prietà di comportamento sforzo-deforma-zione notevolmente simili a quelle dellagomma: essa infatti può essere stirata an-che fino a una lunghezza pari al 170 percento della propria lunghezza originariaallo stato di riposo senza deformarsi inmaniera irreversibile.
La struttura a rete del tessuto connetti-vo può fornire una certa quantità della for-za di recupero che permette alle singolecellule muscolari di ritornare alla proprialunghezza originaria dopo una contrazio-ne. Quando la cellula si contrae e si ispes-sisce, esercita una tensione sulla trama di
fibre che la avvolge. Dopo la contrazione,le fibre possono comprimerla entro un dia-metro inferiore, facendola allungare.
Una trama analoga di fibre di collagenocirconda interamente i muscoli (raggrup-pamenti di numerose cellule muscolari).Questa struttura, anch'essa come una reteper amache, è costituita soprattutto da fi-bre inestensibili, ma ciononostante può es-
sere tirata senza alcun danno in misuraconsiderevole grazie allo schema struttu-rale della sua trama: i trefoli si incrocianoformando parallelogrammi che possonoallungarsi lungo la diagonale maggiore fi-no a quando tutti e quattro i lati siano pres-soché paralleli.
Abbiamo scoperto che quando dellestrisce di muscolo cardiaco isolato di ratto
Il tessuto connettivo può favorire in due modi la tendenza del cuore a dilatarsi attivamente durantela diastole. I trefoli di fibre connettive che avvolgono le singole cellule (in alto) girano in senso siaorario, sia antiorario, formando una rete intrecciata, non dissimile da quella che si utilizza per leamache (a). Numerosi esperimenti indicano che questa struttura a rete con maglie a forma diparallelogrammi protegge la fibra muscolare da una eccessiva trazione: quando la fibra viene sti-rata (b) le maglie si allungano nella direzione della diagonale del parallelogramma e si oppongonoa un ulteriore stiramento. Analogamente, quando il muscolo è totalmente contratto (c), i paralle-logrammi delimitati dalle fibre di tessuto connettivo si stirerebbero nella direzione dell'altra diago-nale, opponendosi forse a un'ulteriore contrazione e tendendo a far estendere di nuovo il muscolo.I montanti di tessuto connettivo che fissano le singole cellule l'una all'altra (in basso) possonoessere sottoposti a tensione e a stiramento (a) quando il cuore si trova in una configurazionequalsiasi diversa da quella naturale di riposo (b). Una situazione di questo tipo può contribuire afar dilatare il cuore dopo che la contrazione sistolica lo ha compresso oltre il suo volume all'equi-librio. Nell'illustrazione sono raffigurate cellule muscolari di due strati adiacenti del miocardio.
6061
In questa microfotografia elettronica a scansione, sottili montanti colle-gano lateralmente cellule muscolari di cuore di mammifero. I montantisono di collageno, una proteina resistente alla trazione, ma contengono
anche una certa quantità di elastina, una proteina estremamente elastica;coordinano il movimento di parti separate del muscolo cardiaco e pos-sono fornire parte della forza che permette al cuore contratto di dilatarsi.
venivano tirate fino a che le fibre di colla-geno che le avvolgevano raggiungevanoquesto limite, la resistenza a un ulterioreallungamento aumentava in modo moltomarcato. Questo è anche il massimo valo-re al quale può essere tirato il muscolo e alquale produce ancora la massima forza.L'interpretazione più semplice e più sugge-stiva di questi risultati è che la trama in-crociata delle fibre di collageno che avvol-ge la cellula muscolare impedisce ai sarco-meri uno stiramento eccessivo. Sembra al-lora ragionevole supporre che tale rete for-nisca anche, quando il muscolo si contraeanziché distendersi, una forza di recuperotale che i parallelogrammi costituiti dallefibre di collageno vengano totalmente este-si lungo l'altra diagonale. Non esiste anco-ra una prova definitiva a sostegno di que-sta ipotesi, anche se la formulazione teori-ca è confortata da una serie di risultatipreliminari ottenuti in diversi laboratori.
C'è un altro modo in cui le fibre di tes-suto connettivo potrebbero favorire
la tendenza del cuore a creare un'aspira-zione dilatandosi attivamente nel corsodella diastole. Nel cuore di mammifero lesingole cellule muscolari sono collegatel'una all'altra lateralmente da elementi si-mili a cordoni, o montanti costituiti so-prattutto da collageno. I montanti appaio-no ritorti a elica, proprio come lo sono icavi di sostegno dei ponti sospesi per offri-re una maggiore resistenza alla trazione.Sono forse essi in grado di fungere da ca-tene, opponendo resistenza alla deforma-zione del cuore e immagazzinando unaparte dell'energia di deformazione elasticaper sfruttarla successivamente nella se-guente fase di aspirazione diastolica?
Un modo per determinare se i montanticontribuiscono all'aspirazione diastolicaconsiste nel mettere a confronto il funzio-namento dei muscoli del cuore di mammi-fero, in cui le cellule muscolari sono inter-connesse da montanti, con il funziona-mento del cuore di anfibio, in cui le cellulemuscolari non sono interconnesse (sebbe-ne, in realtà, le singole cellule siano avvolteda fibre di collageno).
Se il cuore funzionante di un ratto vieneimmerso in una soluzione fisiologica, con-tinua a battere, anche se più lentamentedel normale e si sposta rapidamente nelliquido. Questo viene espulso energica-mente attraverso i grandi vasi durante lasistole e quindi nuovamente aspirato du-rante la diastole. L'esperimento dimostrain modo semplice che, anche nel cuore diratto, esiste una significativa aspirazionediastolica, dal momento che nessuna pres-sione esterna di riempimento spinge il li-quido nel cuore. Se un esperimento analo-go viene eseguito sul cuore di una rana,questo batte molto debolmente e si spostasolo di poco o nulla attraverso il liquido.Una possibile spiegazione è che, nel cuoredi rana, l'aspirazione diastolica non è suf-ficiente ,a far affluire in quantità liquidonelle cavità cardiache e la corrispondenteespulsione del liquido non è sufficiente-mente energica da far spostare il cuore nel-
la direzione opposta. L'ipotesi che il cuoredi ratto abbia un'azione di pompaggio piùvigorosa e un grado di aspirazione mag-giore in parte grazie ai montanti che con-nettono le singole cellule muscolari e coor-dinano la contrazione di gruppi di cellule,è indubbiamente molto suggestiva.
Essa acquista poi ulteriore credibilitàsulla base di studi comparabili che abbia-mo eseguito su due invertebrati. Uno diquesti, il calamaro, aspira l'acqua entrouna cavità centrale, che è in pratica unapompa muscolare, e poi la espelle energi-camente contraendo la cavità stessa e pro-cedendo cosi con un tipo di propulsione areazione. La cavità muscolare del calama-ro è rivestita da un mantello ricco di col-lageno e fissato alle singole fibre muscolarimediante elementi a montante, che ricor-dano i montanti che connettono l'una al-l'altra le singole fibre muscolari nel cuoredi ratto.
La struttura muscolare del polpo, d'al-tro canto, è simile a quella del cuore dirana. Nel polpo ciascuna cellula muscola-re è circondata da fibre di tessuto connet-tivo sinuose, ma non vi è traccia di con-nessioni intercellulari. Il polpo, al contra-rio del calamaro, di solito non procede nel-l'acqua per propulsione a reazione ma sisposta mediante i tentacoli, con movimen-ti sinuosi tra le rocce del fondo marino.(Solo occasionalmente - per darsi alla fugain caso di pericolo - contrae alcune sezionidella propria cavità interna e produce cosìun piccolo effetto a reazione.)
Sulla base di studi preliminari condottisul calamaro, sembra che il tessuto con-nettivo intercellulare del cuore di mammi-fero e del calamaro aumenti l'efficienza diuna contrazione e della successiva aspira-zione, che serve a riempire una cavità do-po che da questa è stato espulso liquido.Sia il cuore di ratto, sia la cavità centraledel calamaro possiedono queste intercon-nessioni e sono entrambi in grado di gene-rare un'aspirazione e di spostarsi mediantepropulsione a reazione attraverso un mez-zo liquido; il cuore di rana e quello di pol-po non possiedono, invece, interconnes-sioni del genere e non mostrano né un'a-spirazione significativa né un movimentoper propulsione a reazione degni di nota.
Forse i montanti favoriscono l'aspira-zione coordinando l'azione di numerosecellule muscolari singole: fissandole l'unaall'altra, essi possono opporre resistenzaalla deformazione del cuore quando que-sto si contrae oltre il proprio volume all'e-quilibrio e così, nel cuore totalmente con-tratto, la tensione dei montanti potrebbecontribuire a immagazzinare una partedell'energia della contrazione sistolica.
L'ipotesi che il tessuto connettivo abbiaun ruolo nella dilatazione diastolica delcuore trova sostegno negli esperimenti cheabbiamo eseguito con Joseph M. C apassodell'Albert Einstein College of Medicine.Abbiamo scoperto che il rapporto tra lavelocità di contrazione e la velocità di ri-distensione è molto minore in un muscolopapillare (muscolo che controlla il movi-mento della valvola mitrale) intero che non
in cellule muscolari isolate. Ciò significache la cellula torna più rapidamente allapropria lunghezza iniziale quando è partedi un tessuto che non quando è isolata dal-le altre cellule e dal tessuto connettivo chele tiene insieme. Noi attribuiamo la diffe-renza tra questi rapporti agli effetti che iltessuto connettivo impone alla dinamicadella contrazione e dell'allungamento dellecellule muscolari.
I 'aspirazione non è l'unico modo in cuil'energia di una contrazione sistolica
può essere applicata alla successiva dila-tazione diastolica. Oltre a modificare laforma del cuore (e la forma delle fibre mu-scolari cardiache), la contrazione cambiala posizione del cuore. I principali vasisanguigni che riforniscono il cuore umanoe quelli che partono da esso sono connessia quell'ampia superficie che è nota comebase del cuore (pur trovandosi, in realtà,superiormente, cioè nel punto più vicino alcapo). Come un calamaro nell'acqua o uncuore di ratto, isolato, immerso in soluzio-ne fisiologica, quando si contrae, spingen-do il sangue verso l'alto, il cuore umanoper reazione si sposta verso il basso, nelladirezione opposta a quella del liquidoespulso. Secondo noi, una certa quantitàdell'energia cinetica che il cuore accumuladurante la sistole viene successivamenteutilizzata per riempire i ventricoli durantela diastole.
Diverse sono le tecniche che permetto-no di studiare il movimento globale delcuore nel torace. La cardiografia balistica,una tecnica sorpassata, prevedeva che ilsoggetto venisse fatto sdraiare su un tavo-lo a ruote; venivano quindi registrati i mo-vimenti minimi del tavolo in risposta allevariazioni della quantità di moto del cuoredi quel soggetto. Con una tecnica più mo-derna, l'ecocardiografia bidimensionale,viene generata un'immagine del cuore inmovimento e in tempo reale utilizzando leonde sonore riflesse a frequenze ultrasoni-che dalla superficie del cuore. Si può anchemisurare la velocità e la direzione con cuiil sangue scorre in determinate regioni delcorpo, focalizzando le onde sonore su que-ste zone e quindi registrando lo sposta-mento Doppler delle onde riflesse.
Le tecniche citate hanno dimostrato cheil cuore compie movimenti sostanziali neltorace. Durante la sistole la sua base siallontana dal capo, sottoponendo a unostiramento sia il tessuto connettivo cede-vole, sia i vasi sanguigni che lo tengono insede. Questi elementi di connessione tra-sformano una frazione dell'energia cineti-ca del cuore in energia potenziale di stira-mento ed esercitano sulla base una forzadiretta verso l'alto. Così la base, durantela diastole, viene nuovamente rinforzata indirezione del capo. Nello spostarsi verso ilsangue in arrivo, sotto l'influsso della for-za elastica di rinculo, il cuore, che si starapidamente dilatando, inghiotte il sanguein arrivo riempiendosi rapidamente.
Il rinculo contribuisce in larga misura alprecoce e accelerato riempimento del cuo-re necessario per un funzionamento effi-
ciente a frequenze cardiache elevate; essopermette, inoltre, riempimenti e svuota-
* menti a basse pressioni e aumenta l'effi-cienza complessiva del meccanismo dipompaggio sfruttando l'energia della si-stole per alimentare la diastole.
Il movimento globale del cuore favori-sce il riempimento della cavità anche in unaltro modo meno appariscente. Il ventri-colo destro, la cui parete è relativamentesottile, è conformato come una sacca cheingloba parzialmente il ventricolo sinistro,così come una mano si flette afferrando unpugno chiuso. L'energico rinculo del ven-tricolo sinistro che si rilascia può in realtàinnalzare il ventricolo destro al di sopradel livello del sangue nell'atrio destro. Ilsangue, che durante la sistole precedenteera fluito nell'atrio destro, deve soltantorimanere relativamente immobile perchél'atrio possa essere circondato dal ventri-colo destro che si sposta. Grazie a questomeccanismo, l'aumentata contrattilità del-la metà sinistra del cuore può incrementa-
re la gettata della metà destra: quando au-menta la contrattilità del ventricolo sini-stro, questo subirà un maggiore svuota-mento sistolico, un rinculo maggiore espingerà con una forza maggiore il ventri-colo destro oltre il livello del sangue con-tenuto nell'atrio destro.
Nel modello che proponiamo, il riempi-mento precoce del ventricolo avviene
sotto il controllo del rilasciamento del mu-scolo cardiaco e della dilatazione della ca-vità, oltre che del rinculo balistico del cuo-re. Il volume finale del ventricolo è quindiregolato dalla pressione del sangue in ar-rivo. In alcuni stati patologici il cuore hameno rinculo e capacità di aspirazione e siriempie in modo incompleto. In situazionidi questo tipo, la legge di Frank-Starlingconvenzionale descrive con accuratezzala funzione cardiovascolare. Le pressioniesercitate dal riempimento diastolico au-mentano allo scopo di accrescere il volumedel sangue espulso con il battito successi-
vo, ma pressioni superiori possono ancheprovocare una congestione venosa, chepuò dar luogo, alla fine, a un'insufficienzacardiaca congestizia. Nel nostro modelloil cuore è visto come una pompa ad aspi-razione dinamica che dipende per il pro-prio funzionamento dalle proprietà coor-dinate e integrate di una gerarchia di strut-ture a tutti i livelli di dettaglio. I livelli van-no dalla riespansione della cellula musco-lare causata da elementi citoscheletrici eda doppie sovrapposizioni all'interno deisarcomeri, al movimento globale del cuorenel torace attraverso la tensione del tessu-to connettivo che avvolge le fibre musco-lari e lega una cellula all'altra. Il movimen-to del cuore viene visto come un sistemaunificato, in cui cialcuna fase contribuisceal funzionamento efficiente anche della fa-se successiva. Ulteriormente perfezionato,questo modello potrà fornire una spiega-zione completa della funzione cardiaca,una spiegazione che includa il modello diFrank-Starling come caso limite.