56
Enrico Pieroni [email protected] 27 Maggio 2015 Modellistica molecolare e applicazioni alla sclerosi multipla
| Date post: | 12-Aug-2015 |
| Category: |
Science |
| Upload: | crs4-research-center-in-sardinia |
| View: | 42 times |
| Download: | 2 times |

Enrico Pieroni
27 Maggio 2015
Modellistica molecolare e applicazioni alla sclerosi multipla

Outline
Cos'è un modello? Cos'è una simulazione? L'importanza delle scale temporali e spaziali Il riconoscimento antigenico La sclerosi multipla Il mimetismo molecolare Riconoscimento molecolare in sclerosi multipla Risultati delle simulazioni sulle proteine collegate alla
sclerosi multipla
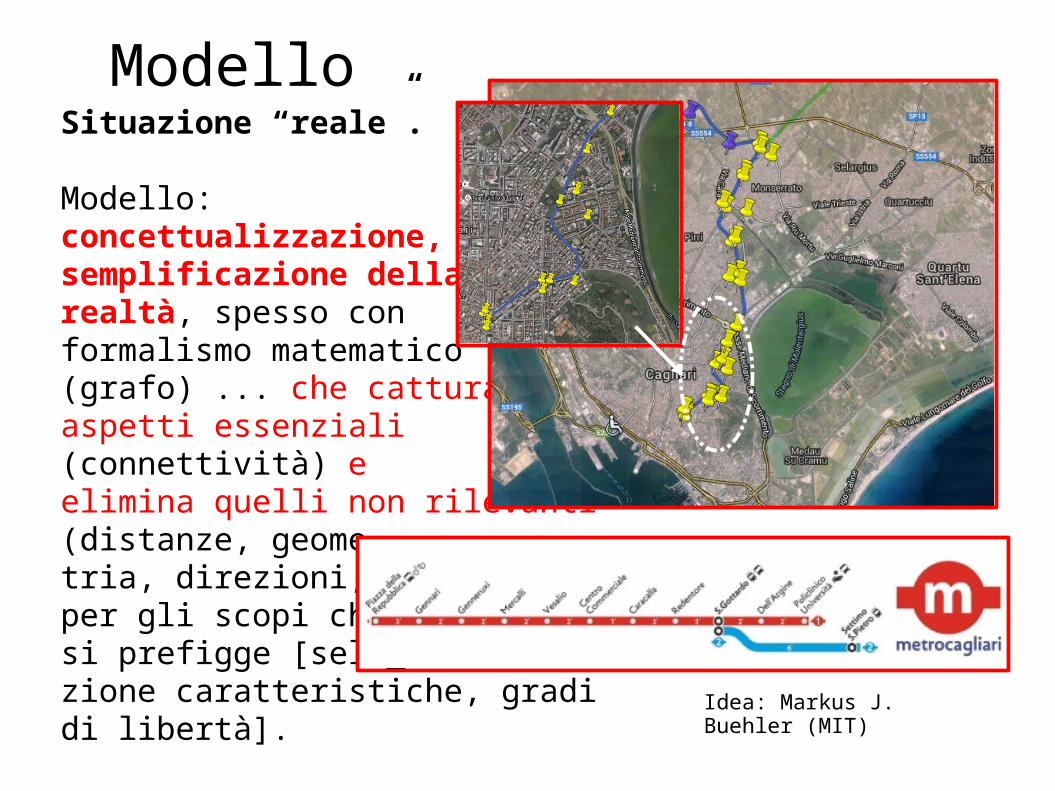
ModelloSituazione “reale”.
Modello:concettualizzazione, semplificazione della realtà, spesso conformalismo matematico (grafo) ... che cattura aspetti essenziali (connettività) eelimina quelli non rilevanti(distanze, geome_tria, direzioni, ...)per gli scopi che ci si prefigge [sele_zione caratteristiche, gradi di libertà]. Idea: Markus J. Buehler (MIT)

Simulazione
Cosa succederebbe se cavalcassi un raggio di luce? (A. Einstein, 1905)

SimulazioneSoluzione (delle equazioni) del modello.
Rappresenta una sorta di esperimento virtuale.
Perchè simulare? Gli esperimenti talvolta sono impossibili, costosi, pericolosi, richiedono tempi lunghi, alcune scale temporali e spaziali di interesse non sono accessibili, ...
Inoltre, la simulazione consente di: interpretare i dati “sperimentali”, aiutare a capire i meccanismi sottostanti e predire nuove proprietà del sistema reale, suggerire/selezionare esperimenti, aiutarne la progettazione, ...
La simulazione offre un quadro dinamico, dove è possibile cambiare i parametri del sistema alla ricerca di proprietà emergenti (sistemiche).

Simulazione al CalcolatoreOggi la simulazione avviene spesso tramite computer, ... e
la velocità di simulazione scala con la potenza computazionale!
Per cui oggi è possibile simulare sistemi complessi, quindi interessanti, quali i sistemi biologici.
Blue Gene 2011, circa 700 TeraFLOPS: 700 000 000 000 000 op./s1 giorno
ENIAC 1946, 5 000 operazioni/scirca 2.5 milioni di secoli

Sistemi a diverse scale L-T
Elaborazione da idea di Aatto Laaksonen (Stokholm U.)
ps
ns
μs
fs
m = 1 / 1 000μ = 1 / 1 000 000n = 1 / 1 000 000 000p = 1 / 1 000 000 000 000f = 1 / 1 000 000 000 000 000
psps
1 Å
Cristallografia a raggi X, spettroscopia NMR,
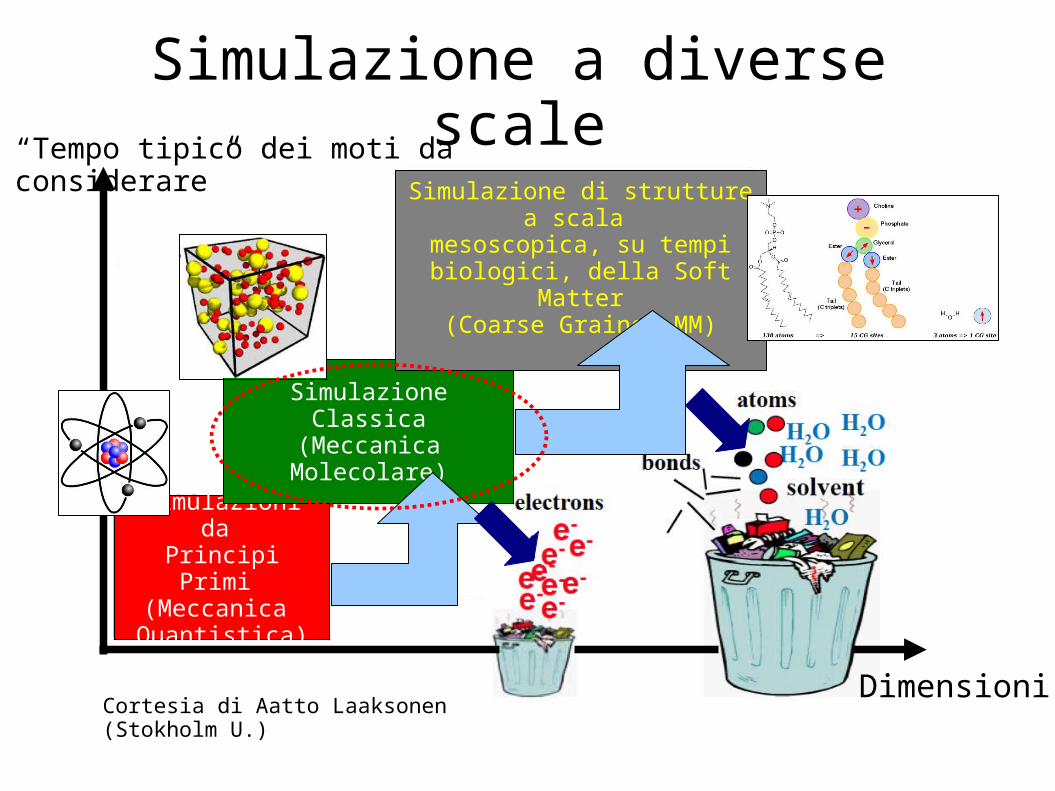
Simulazione a diverse scale
Simulazioni da Principi Primi (Meccanica Quantistica)
Simulazione Classica(Meccanica Molecolare)
Simulazione di strutture a scala mesoscopica, su tempi
biologici, della Soft Matter(Coarse Grained MM)
Cortesia di Aatto Laaksonen (Stokholm U.)Dimensioni
“Tempo tipico dei moti da considerare”

μicro – Macroscopico
Simulazione molecolare: genera tante configurazioni del sistema, misura le proprietà statistiche e (tramite la fisica statistica) le collega a quelle macroscopiche
Alcuni parametri importanti: Scala temporale dei moti inclusi nel modello (condiziona il passo
di risoluzione delle equazioni)
Scala temporale degli effetti osservabili di interesse su scala macroscopica (condiziona il tempo totale di simulazione)
Dimensione del sistema (condiziona il tempo totale di simulazione)
Comportamento μ:moto atomico e
molecolare, forze interatomiche ed
intermolecolari
Comportamenti M osservabili:proprietà termodinamiche (T,P,V),di trasporto (velocità di diffusione),
ottiche, ...
Comportamento M bio:Legame, modifiche strutturali,
attivazione recettore,diffusione farmaci
attraverso membrana, ...

Applicazioni della Modellistica Molecolare
Struttura delle proteine, per omologia con strutture cristallografiche.
Interazione Proteina:Molecola, ad esempio Farmaco:Recettore.
Interazione Proteina:Peptide, ad esempio in segnalazione e regolazione cellulare, riconoscimento degli antigeni (p:MHC).
Proteina:Proteina ad es. pMHC:TCR, metabolismo, segnalazione cellulare, funzionamento muscolare.
Proteina:Lipide, ad es. Recettore:Lipide, segnalazazione cellulare, sistema immunitario, sindrome metabolica.
Proteina:Carboidrato, ad es. in infiammazioni, riconoscimento patogeni, adesione e segnalamento cellulare.
Proteina:Polimero, ad esempio per il trasporto dei farmaci.
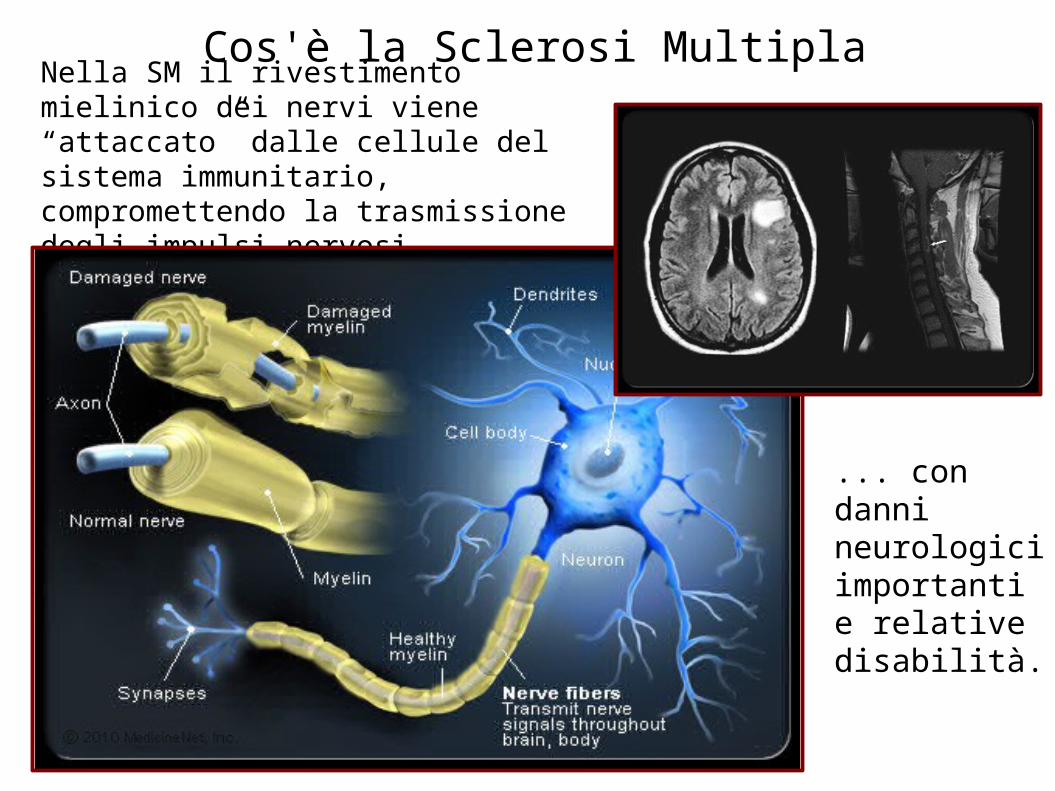
Nella SM il rivestimento mielinico dei nervi viene “attaccato” dalle cellule del sistema immunitario, compromettendo la trasmissione degli impulsi nervosi ...
... con danni neurologici importanti e relative disabilità.
Cos'è la Sclerosi Multipla

Fattori Genetici
Evidenza:Gemelli monozigotici: 25-30%Gemelli dizigotici: 3-4%
MHC (HLA): principale determinante genetica
Quali sono le cause?
Fattori Ambientali(sconosciuti!)
-Esposizione solare (Vit. D),-Agenti patogeni, part. virus di Epstein-Barr (EBV),Micobatterio MAP-Fumo.
Cocco, E., Sardu, C., Pieroni, E., et al. 2012 PLoS OneInternational Multiple Sclerosis Genetics, C., Wellcome Trust Case Control, Sawcer, S.,et al. 2011 Nature
Inaugurazione AA 2014-2015 Prolusione della Prof.ssa MG Marrosu, “Immunità in Sardegna: dono degli dei o mela avvelenata?” http://www.unica.it/UserFiles/File/Utenti/verdeoro/2014/12/prolusione2014_marrosu.pdf
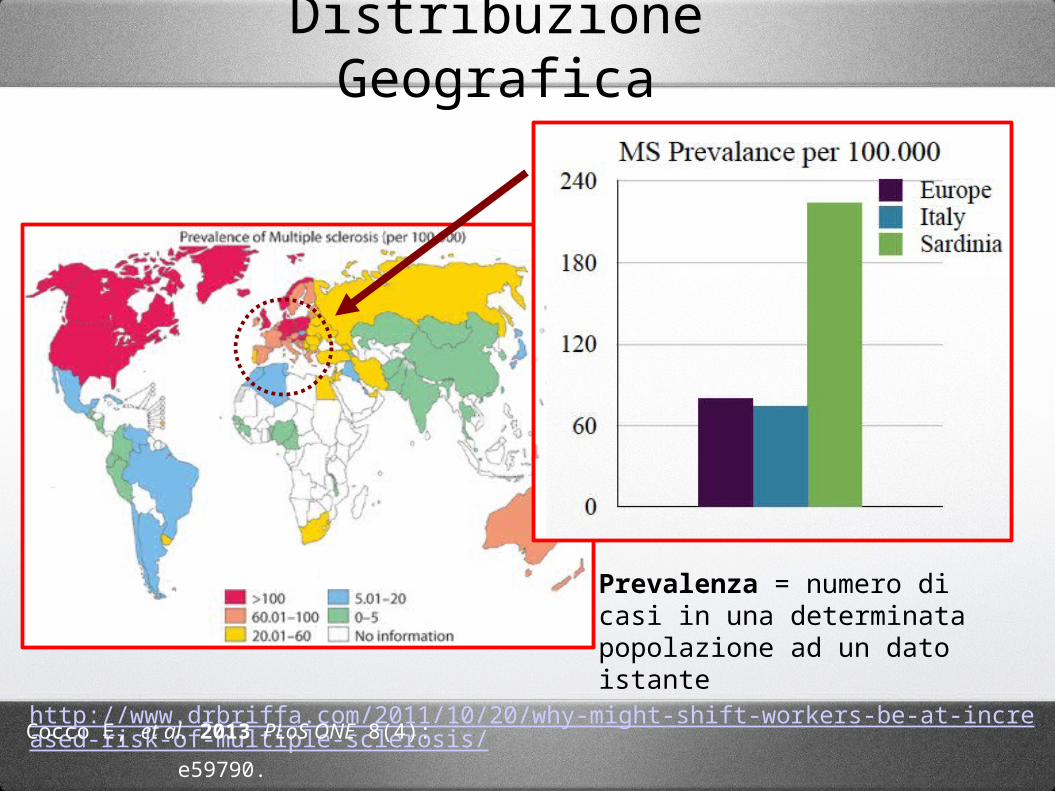
http://www.drbriffa.com/2011/10/20/why-might-shift-workers-be-at-increased-risk-of-multiple-sclerosis/Cocco E, et al. 2013 PLoS ONE 8(4): e59790.
Distribuzione Geografica
Prevalenza = numero di casi in una determinata popolazione ad un dato istante
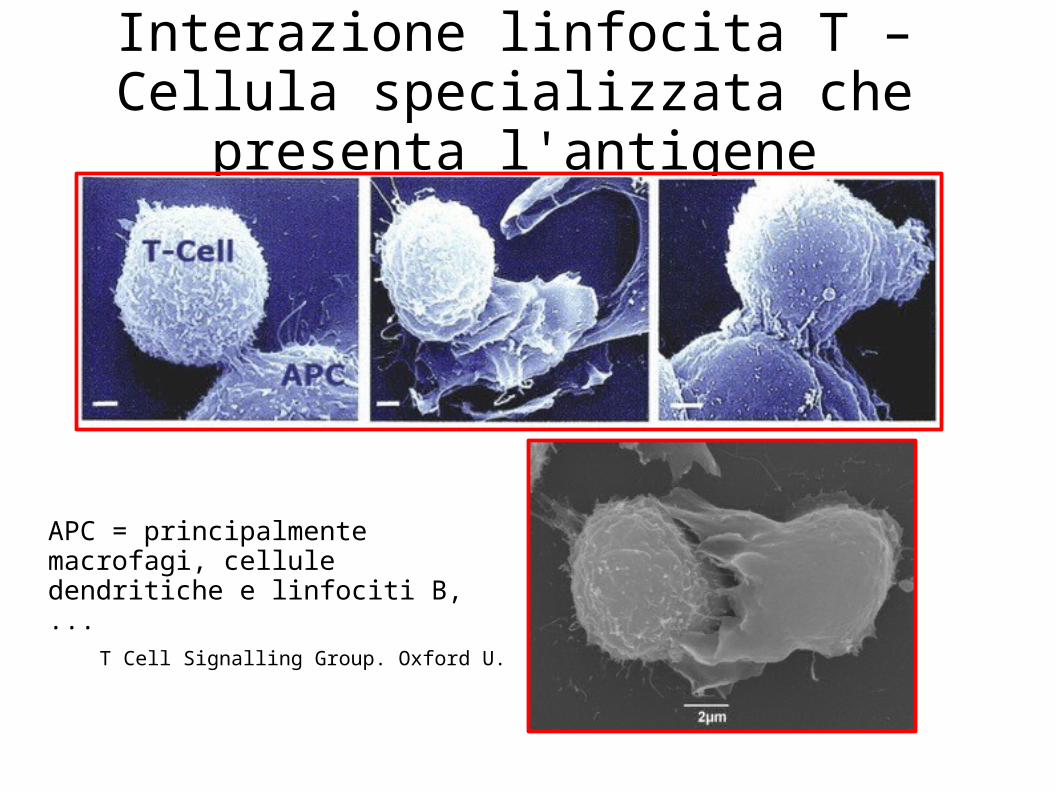
T Cell Signalling Group. Oxford U.
Interazione linfocita T – Cellula specializzata che presenta l'antigene
APC = principalmente macrofagi, cellule dendritiche e linfociti B, ...
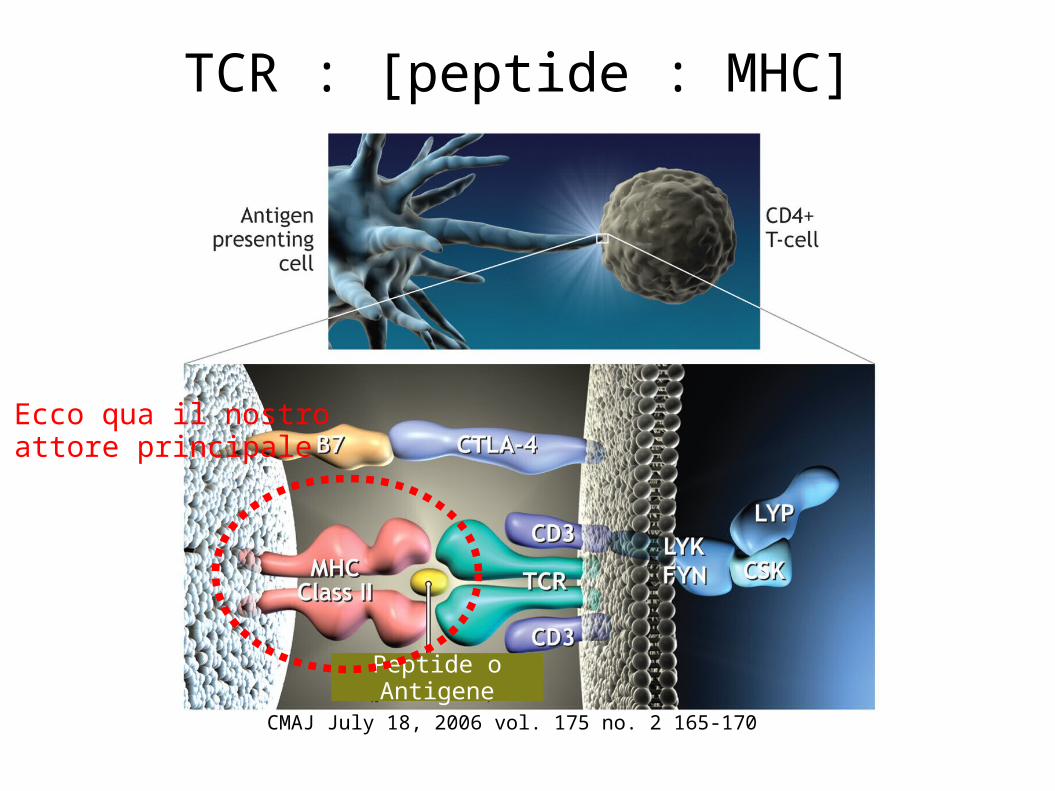
CMAJ July 18, 2006 vol. 175 no. 2 165-170
TCR : [peptide : MHC]
Ecco qua il nostroattore principale
Peptide o Antigene
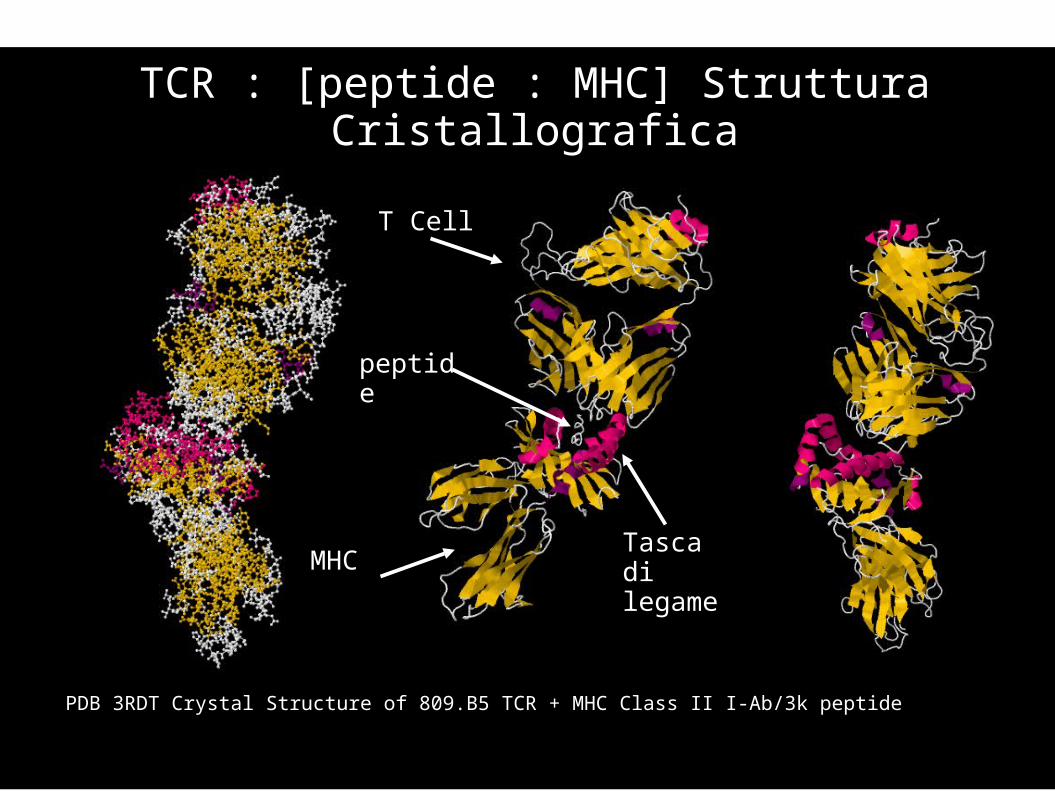
PDB 3RDT Crystal Structure of 809.B5 TCR + MHC Class II I-Ab/3k peptide
TCR : [peptide : MHC] Struttura Cristallografica
T Cell
peptide
MHCTasca di legame
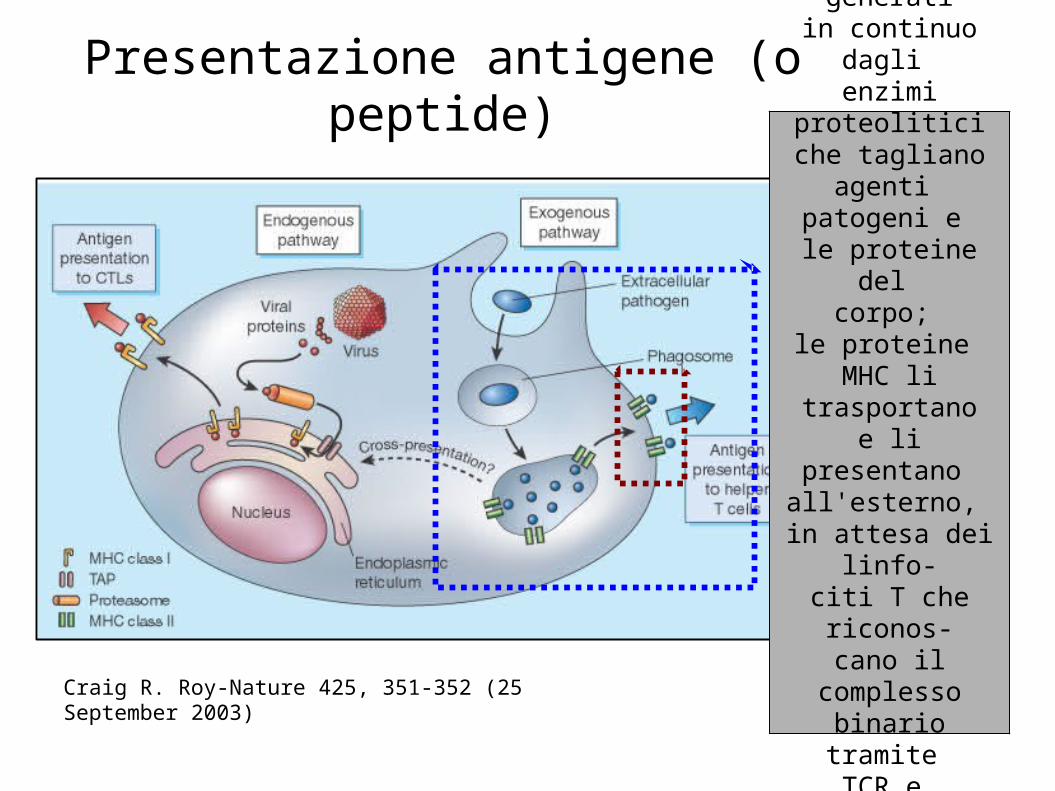
Presentazione antigene (o peptide)
Craig R. Roy-Nature 425, 351-352 (25 September 2003)
Peptidi generatiin continuo
dagli enzimi
proteoliticiche tagliano
agenti patogeni e
le proteine del corpo;
le proteine MHC li
trasportanoe li presentano
all'esterno, in attesa dei
linfo-citi T che riconos-cano il
complessobinario tramite TCR e avviino
la reazione immunitaria.

La Dinamica Molecolare HLA-DRB1:1501
- EBV
La DM è, in pratica, un filmino, con tante informazioni visive sulle configurazioni del complesso, che vanno poi quantificate attraverso l'analisi statistica di tutti i fotogrammi.

Investigare la plasticità del sistema peptide-
MHC, confrontare i risultati per peptidi del
proprio corpo e provenienti da agenti patogeni,
correlare le caratteristiche strutturali e
dinamiche del complesso peptide-MHC alla
suscettibilità verso la malattia, utilizzando
tecniche di Dinamica Molecolare.
Il nostro Obiettivo
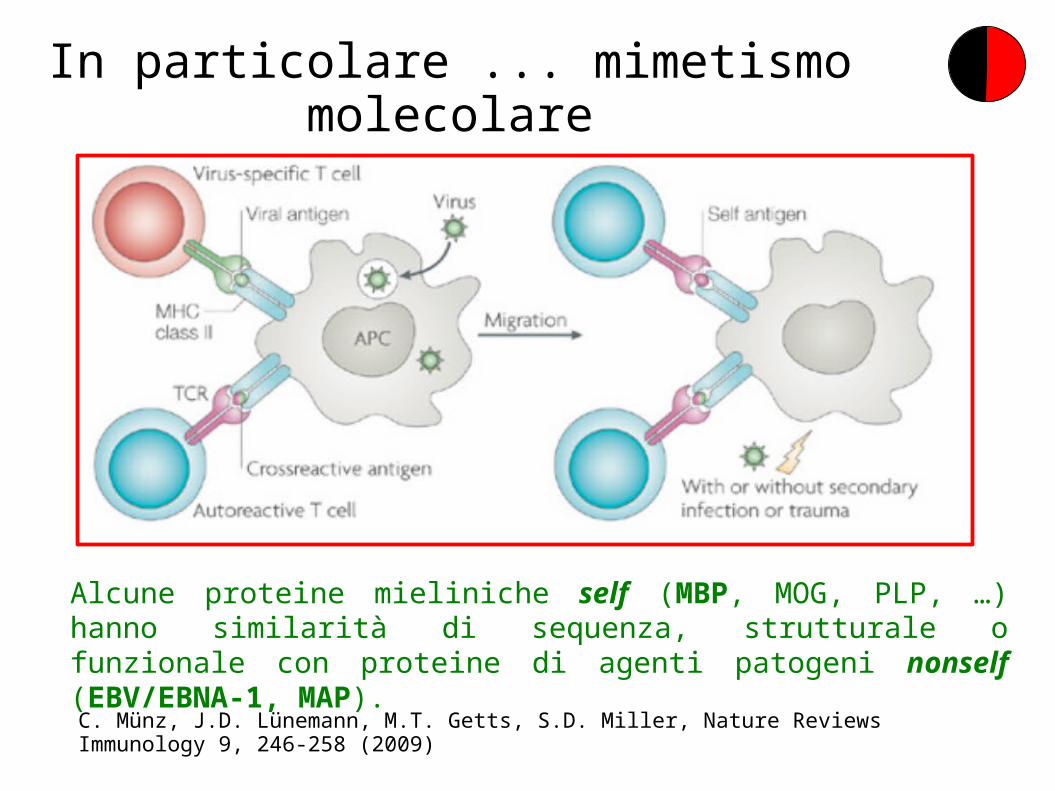
In particolare ... mimetismo molecolare
C. Münz, J.D. Lünemann, M.T. Getts, S.D. Miller, Nature Reviews Immunology 9, 246-258 (2009)
Alcune proteine mieliniche self (MBP, MOG, PLP, …) hanno similarità di sequenza, strutturale o funzionale con proteine di agenti patogeni nonself (EBV/EBNA-1, MAP).

2012
Si parte dalla Genomica

Le prime simulazioni molecolari

RMSD(root mean square deviation)distanza media fra gli atomi fra due strutture sovrapposte(tipicamente tali atomi sono Cα del backbone-ossatura, e la sovrapposizione è ottenuta in maniera rigida ed ottimale)
In un filmino di DM, le due strutture sono quella iniziale (frame a t=0) e quella ad un certo istante (frame al tempo t)
Distribuzione (Istogramma)Quante volte un dato valore è stato osservato in un frame del filmino generato dalla Dinamica Molecolare.Negli esempi successivi: RMSD e larghezza della tasca di legame
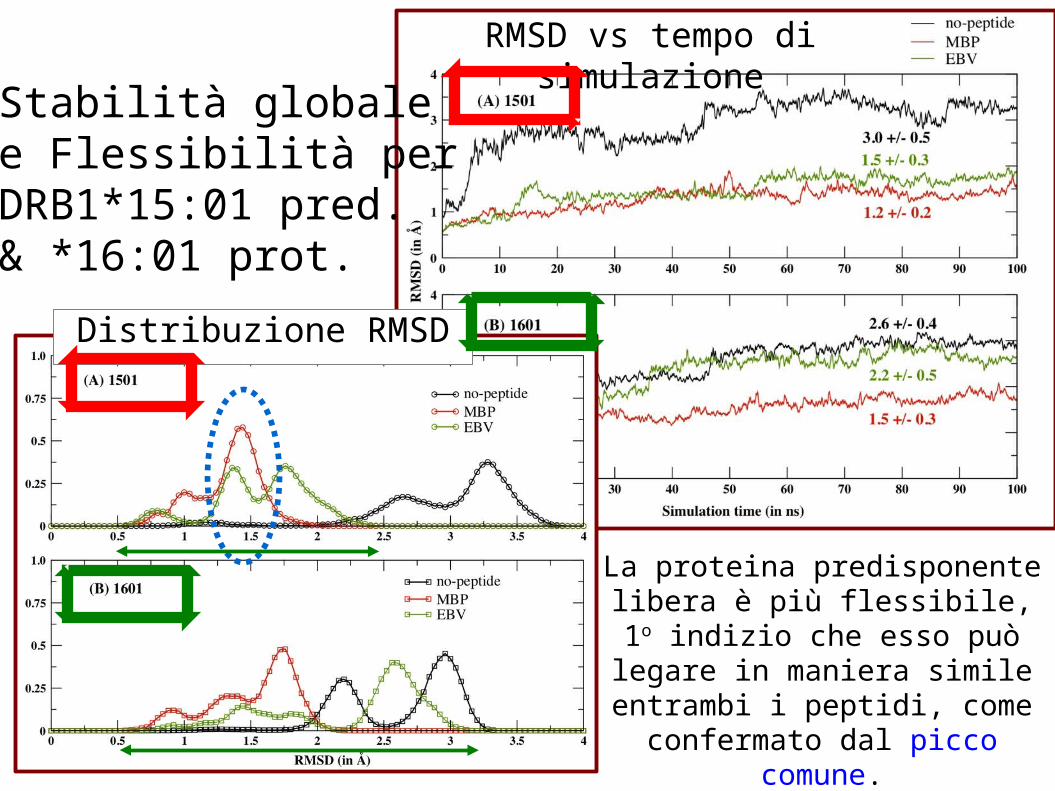
Stabilità globalee Flessibilità perDRB1*15:01 pred.& *16:01 prot.
La proteina predisponente libera è più flessibile, 1o
indizio che esso può legare in maniera simile entrambi i
peptidi, come confermato dal picco comune.
RMSD vs tempo di simulazione
Distribuzione RMSD
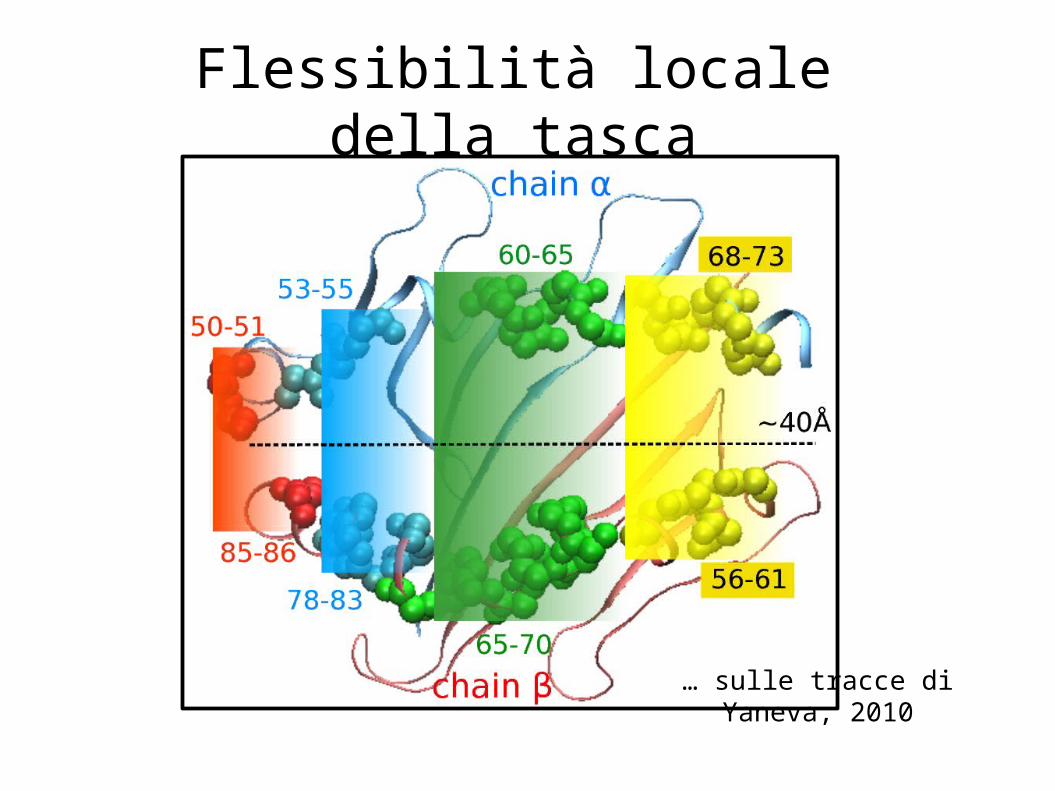
Flessibilità locale della tasca
… sulle tracce di Yaneva, 2010
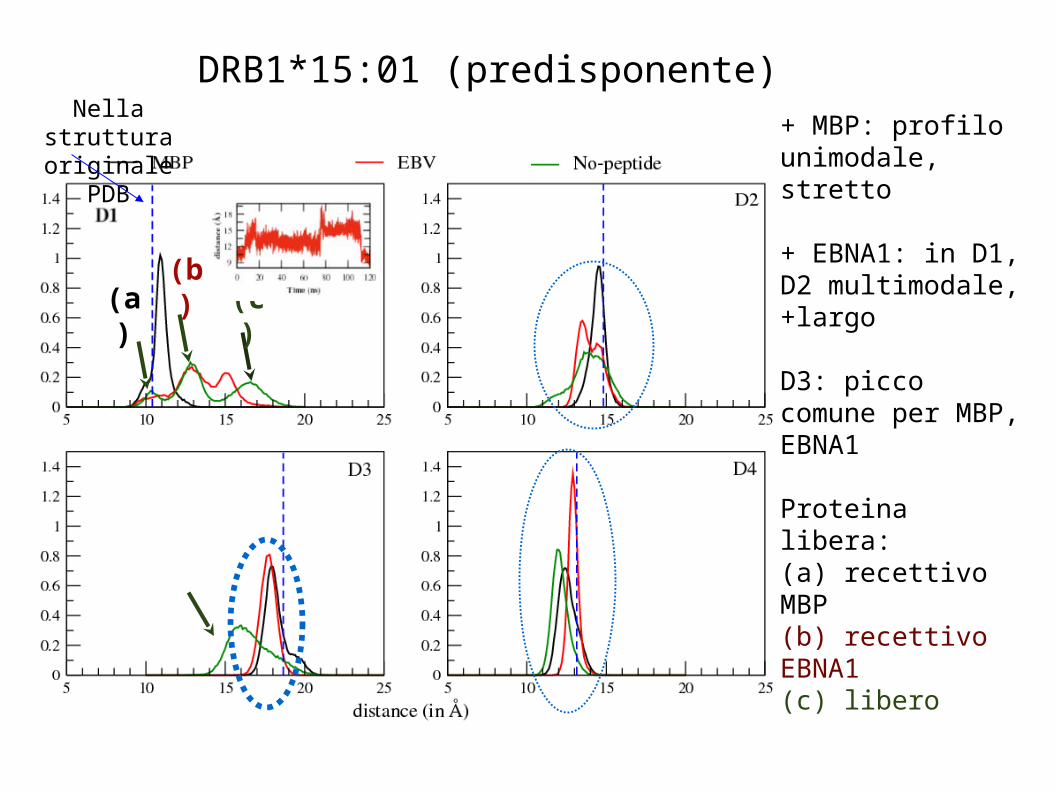
DRB1*15:01 (predisponente)
(a)
(b) (c
)
Nella struttura originale PDB
+ MBP: profilo unimodale, stretto
+ EBNA1: in D1, D2 multimodale, +largo
D3: picco comune per MBP, EBNA1
Proteina libera:(a) recettivo MBP(b) recettivo EBNA1(c) libero
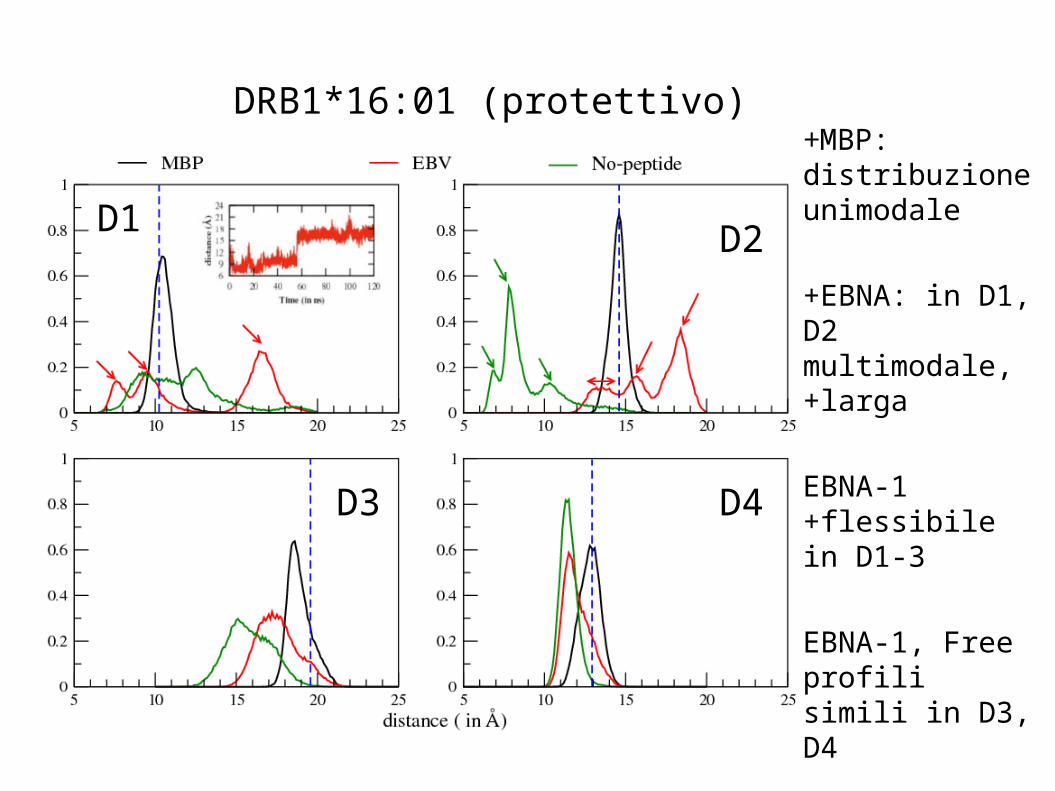
DRB1*16:01 (protettivo) +MBP: distribuzione unimodale
+EBNA: in D1, D2 multimodale, +larga
EBNA-1 +flessibile in D1-3
EBNA-1, Free profili simili in D3, D4
D1 D2
D4D3
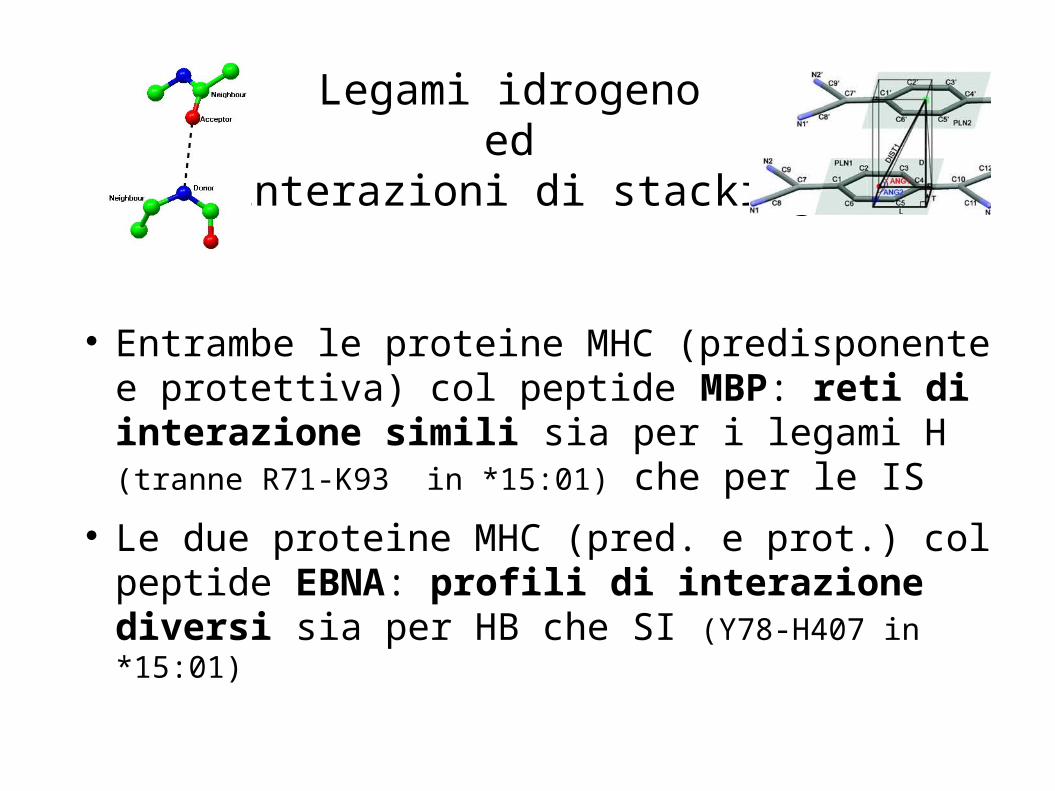
Legami idrogeno ed
interazioni di stacking
Entrambe le proteine MHC (predisponente e protettiva) col peptide MBP: reti di interazione simili sia per i legami H (tranne R71-K93 in *15:01) che per le IS
Le due proteine MHC (pred. e prot.) col peptide EBNA: profili di interazione diversi sia per HB che SI (Y78-H407 in *15:01)
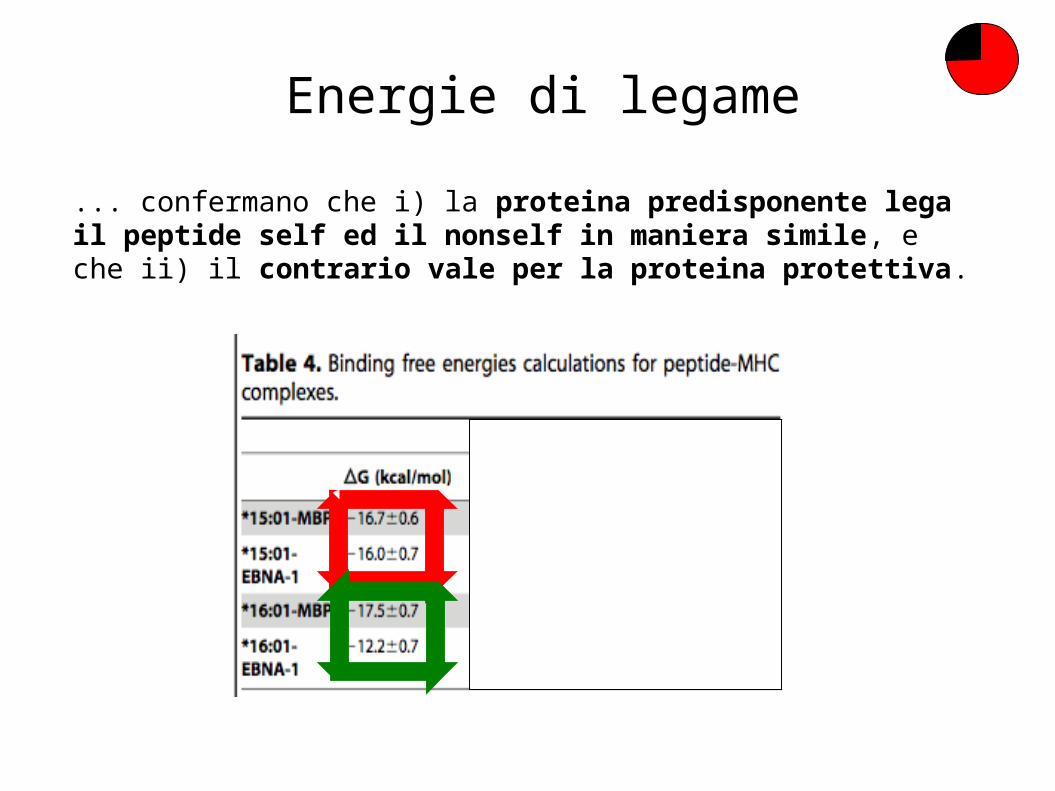
Energie di legame
... confermano che i) la proteina predisponente lega il peptide self ed il nonself in maniera simile, e che ii) il contrario vale per la proteina protettiva.

Primo “take home message” La proteina MHC DR2 predisponente lega in
maniera simile sia il peptide derivato da MBP che quello derivato da EBV, ...
... mentre la proteina protettiva si lega più debolmente al peptide virale e mostra configurazioni di legame nettamente distinte coi due peptidi.
Questo è in accordo con l'ipotesi di mimetismo molecolare: il peptide self (derivato dalla mielina) e quello nonself (virale) hanno somiglianza strutturale e funzionale nel legarsi alla proteina predisponente.

Supporto Sperimentale
I pazienti EBV+ hanno risposta immunitaria anticorpale (mediata dai linfociti B) simile nei confronti dei peptidi EBNA1 ed MBP.
Anticorpi reattivi contro il peptide MBP sono cross-reattivi contro il peptide EBNA1.
EBNA1 ed MBP non hanno omologia di sequenza, quindi devono condividere omologia conformazionale quando sono presentati dalla proteina MHC.

Estensione alle proteine
MH
C “di tipo” D
Q

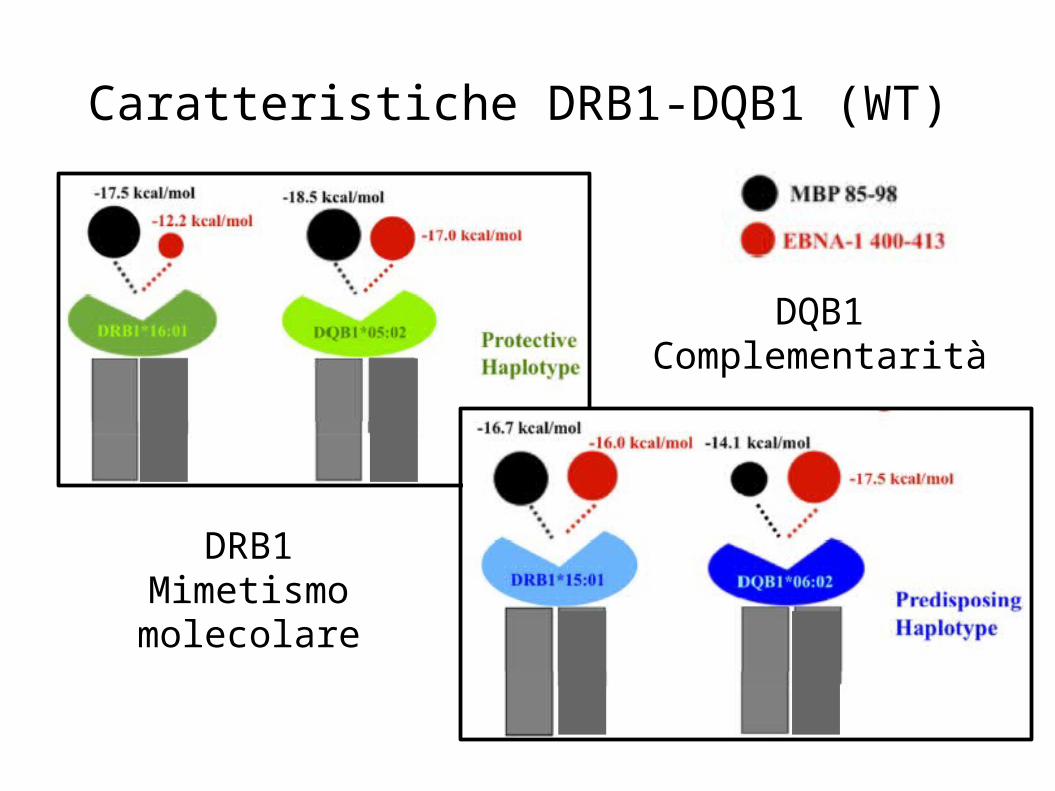
Caratteristiche DRB1-DQB1 (WT)
DRB1Mimetismo molecolare
DQB1Complementarità

Altre analisi ...
Un breve accenno: l'analisi delle reti di interazione idrogeno e di stacking, le distribuzioni di larghezza locale della tasca di legame, sia sulle proteine WT che su quelle mutanti, mostrano i dettagli del processo molecolare di riconoscimento da parte delle proteine DRB1 e DQB1, in particolare della specificità relativamente al tipo di antigene (self/nonself) e di tessuto, e nel loro complesso confermando l'ipotesi di mimetismo molecolare.

Secondo “take home message” APC con proteine protettive presenta complessi con
MBP stabili e duraturi sia attraverso la proteina DRB1 che la proteina DQB1, ...
... e complessi con EBNA1 di vita intermedia solo attraverso la proteina DQB1.
Livelli più alti di presentazione dell'antigeneEfficienza di attivazione superiore (forse) tramite percorsi di attivazione del TCR complementari.
Le proteine predisponenti DRB1 hanno in parte perso l'abilità di discriminazione self/nonself, per cui
la predisposizione verso la SM può essere legata alla perdita della discriminazione self/nonself nel processo di riconoscimento molecolare dell'antigene.

Terzo “take home message”Le proteine DRB e DQB sembrano avere un sistema di
riconoscimento complementare e specifico a seconda del tessuto.
Dato che: i) l'espressione delle proteine DRB1 e DQB1 è dipendente dal tessuto e ii) il mimetismo molecolare richiede una attivazione periferica dei linfociti T prima della riattivazione dei linfociti autoreattivi a livello del sistema nervoso centrale ....
Il ruolo della proteina DRB1 potrebbe essere maggiormente rilevante nella fase di attivazione periferica, ossia di “incontro” iniziale con il fattore scatenante ambientale, mentre ...
... il ruolo della proteina DQB1 potrebbe essere fondamentale nella riattivazione periferica del repertorio di linfociti T clonalmente espansi a partire dal primo incontro, che andrebbero poi “all'attacco” della mielina.

Supporto sperimentale del ruolo DRB/DQB

Supporto sperimentale del ruolo DRB/DQB

Progetto di piccoli peptidi ...
che modulino l'affinità verso peptidi self/nonself,
tramite l'aumento o la soppressione del caricodel peptide nella tasca MHC,
... che agiscano selettivamente rispetto ai tessuti(e quindi alle fasi di arruolamento erroneo dei linfociti T).
Integrazione di dati fenotipici, clinici, medici, genetici, metabolici e di modelistica, al fine di realizzare un modello individuale di rischio nei confronti della malattia e personalizzare il trattamento terapeutico.
Potenzialità in Terapia

Conclusioni
La modellistica computazionale a scala molecolare consente di:Integrare le informazioni genetiche e “sperimentali”;Esplorare potenziali meccanismi funzionali nella SM;<
In prospettiva:Aiutare il progetto di molecole e peptidi terapeutici;Suggerire nuovi “esperimenti”.
Come è tipico per i sistemi biologici, non abbiamo una teoria generale, quanto visto si applica a determinate proteine (pur collegate alla malattia) e determinati peptidi, occorreranno altri esperimenti, dati e simulazioni per ampliare i risultati.


(Modeling) Limits/Perspectives
MS: + alleles, + peptides, TCR;
FMS: less relevant lysoPCs, in general interaction of molecules diet-derived with biological processes;
CASQ2: cardiac isoform;
Therapeutical peptides, tissue- and antigen-selective, modulating autoimmune response;
Molecules interfering with PAFR & modulating pain response;
FMS bio-markers from metabolic profiles:
Clinical and phenotypical data integration;
Individual Risk model, Prognosis and Drug response in MS;
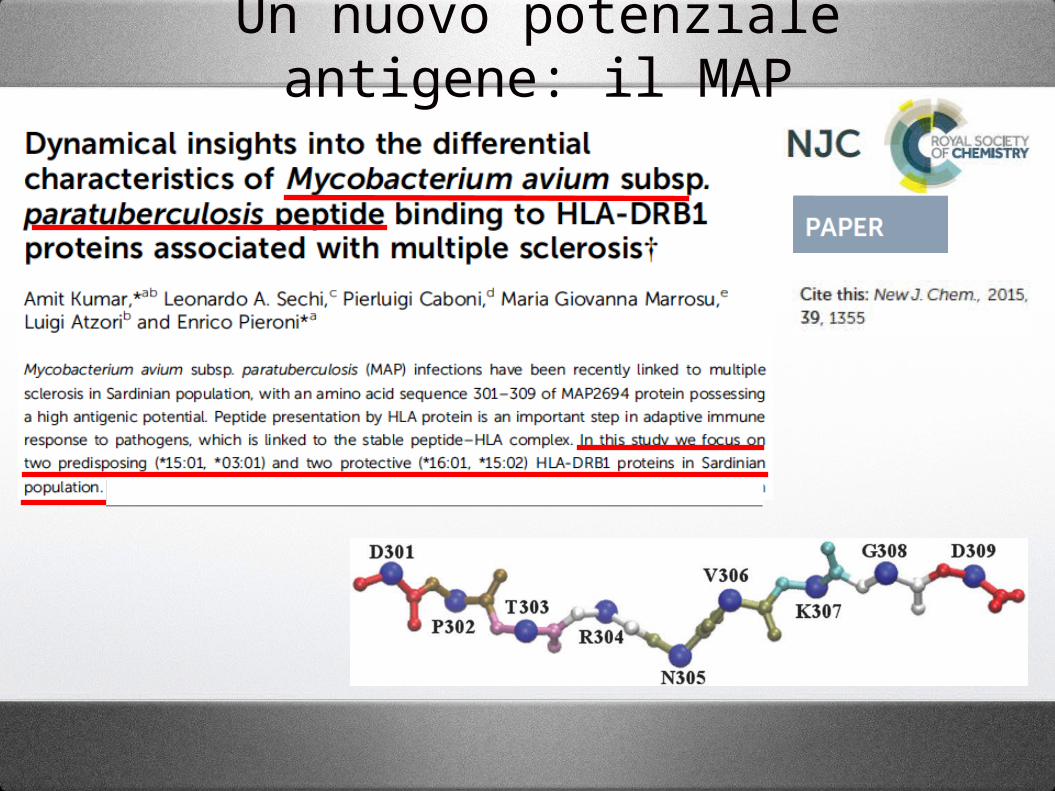
Un nuovo potenziale antigene: il MAP
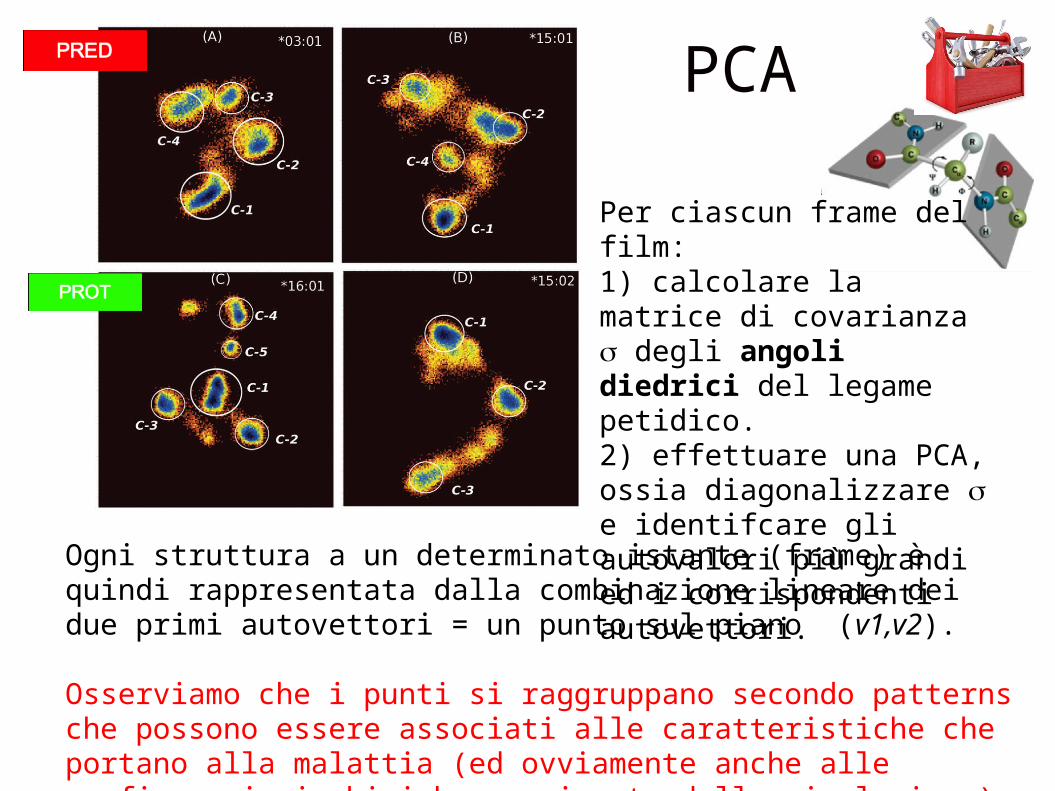
PCA
Ogni struttura a un determinato istante (frame) è quindi rappresentata dalla combinazione lineare dei due primi autovettori = un punto sul piano (v1,v2).
Osserviamo che i punti si raggruppano secondo patterns che possono essere associati alle caratteristiche che portano alla malattia (ed ovviamente anche alle configurazioni chimiche campionate dalla simulazione)
Per ciascun frame del film:
1) calcolare la matrice di covarianza σ degli angoli diedrici del legame petidico.2) effettuare una PCA, ossia diagonalizzare σ e identifcare gli autovalori più grandi ed i corrispondenti autovettori.
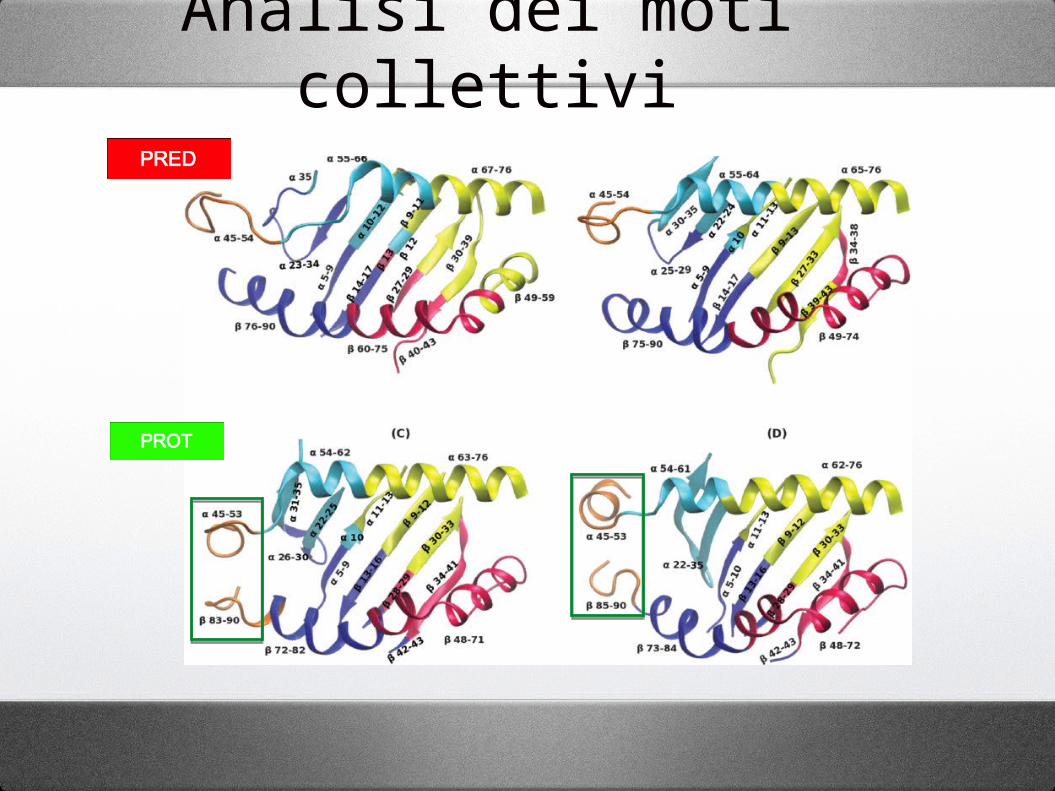
Analisi dei moti collettivi
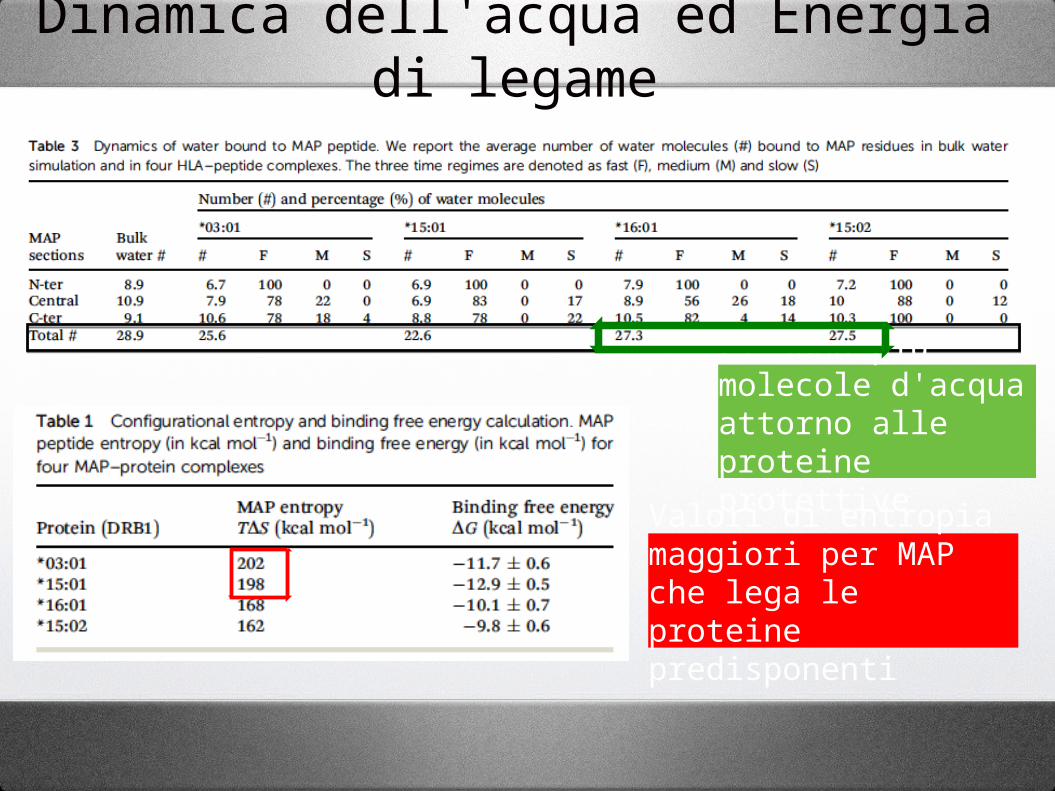
Dinamica dell'acqua ed Energia di legame
Ci sono più molecole d'acqua attorno alle proteine protettive
Valori di entropia maggiori per MAP che lega le proteine predisponenti
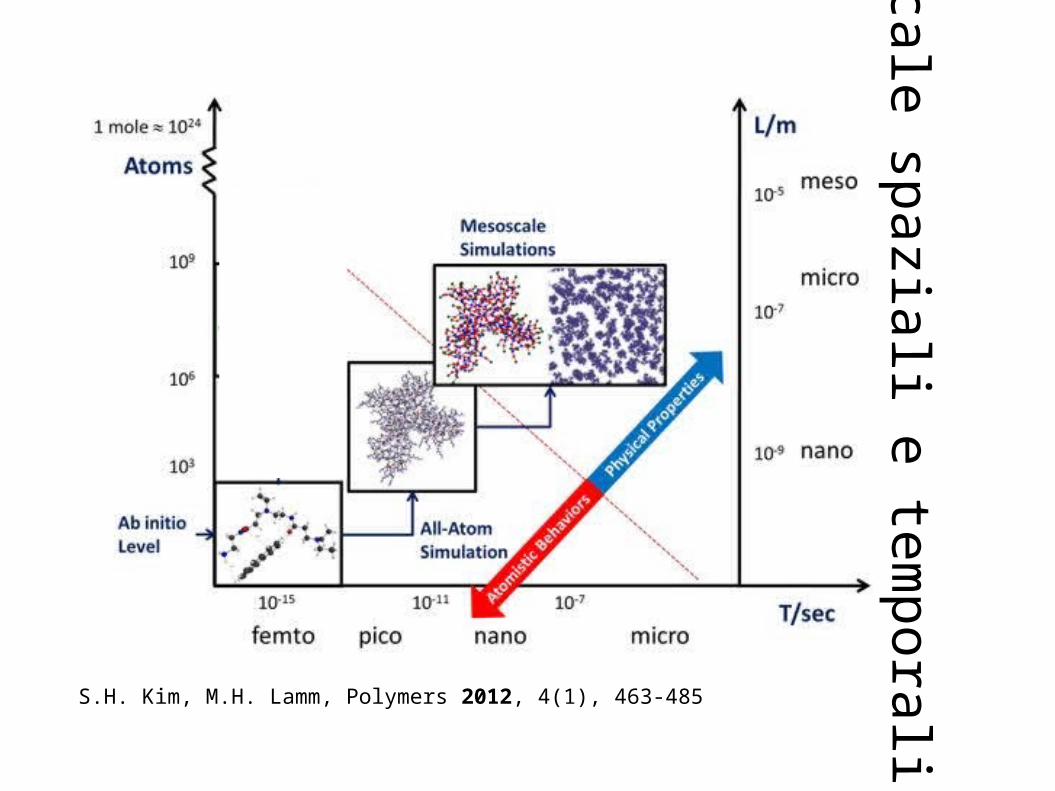
S.H. Kim, M.H. Lamm, Polymers 2012, 4(1), 463-485
Scale spaziali e tem
porali

4
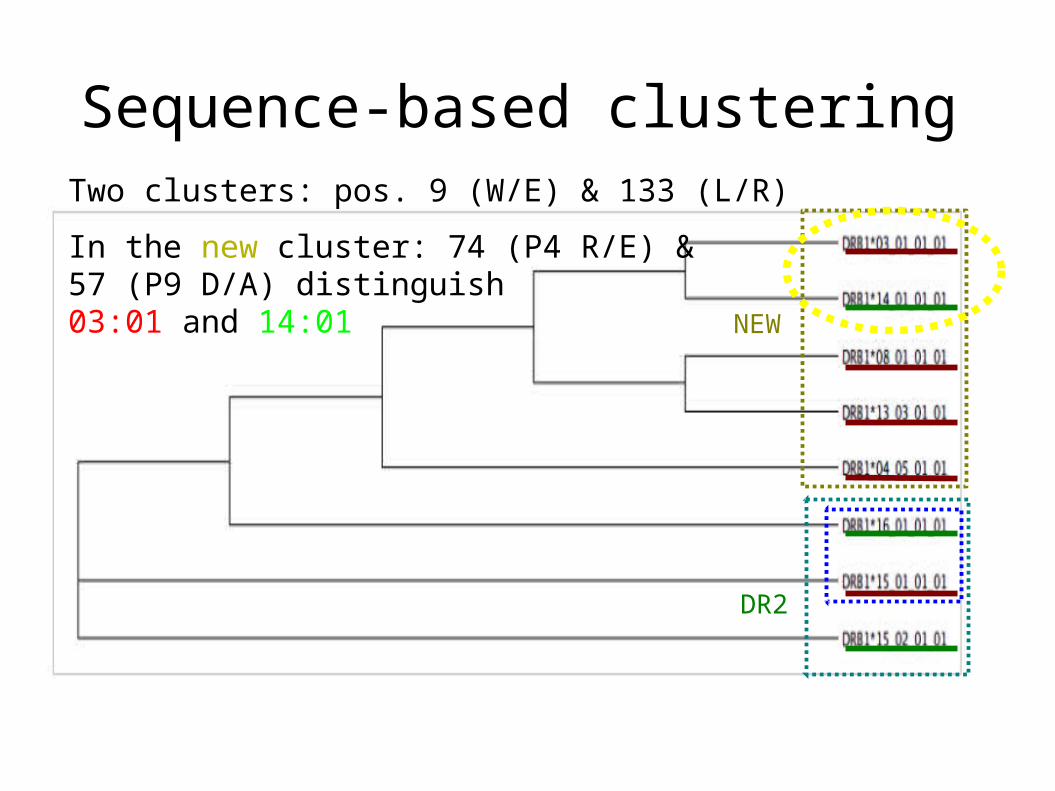
DR2
Sequence-based clusteringTwo clusters: pos. 9 (W/E) & 133 (L/R)
In the new cluster: 74 (P4 R/E) & 57 (P9 D/A) distinguish 03:01 and 14:01 NEW
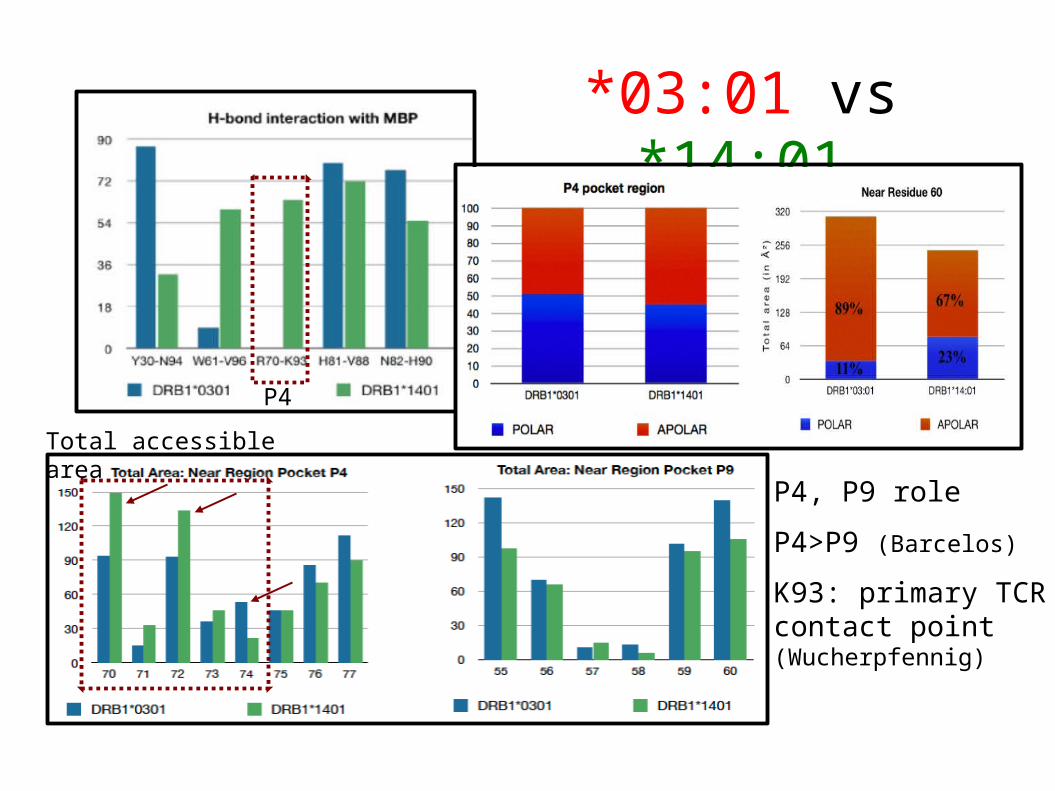
*03:01 vs *14:01
P4
P4, P9 role
P4>P9 (Barcelos)
K93: primary TCR contact point (Wucherpfennig)
Total accessible area
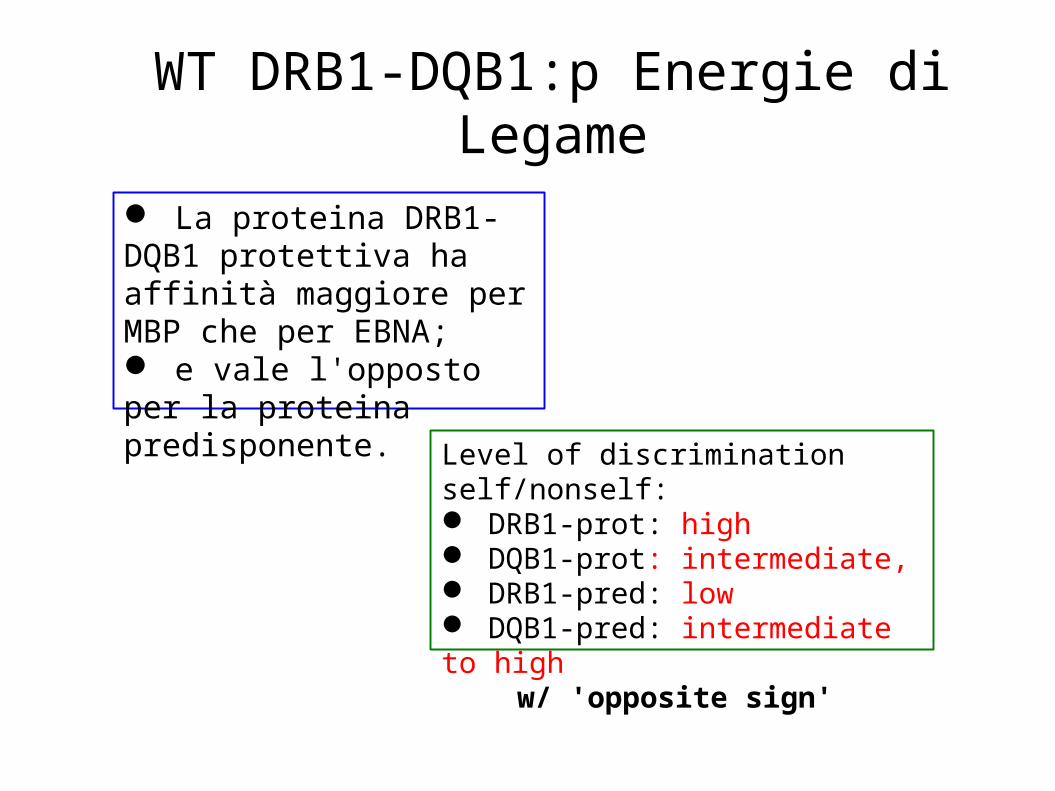
WT DRB1-DQB1:p Energie di Legame
Level of discrimination self/nonself: DRB1-prot: high DQB1-prot: intermediate, DRB1-pred: low DQB1-pred: intermediate to high
w/ 'opposite sign'
La proteina DRB1-DQB1 protettiva ha affinità maggiore per MBP che per EBNA; e vale l'opposto per la proteina predisponente.

Discussion
*03:01, *04:01: very similar by sequence but striking functional differences, particularly binding MBP.
Synergistic interactive effect: resistance allele *14:01 binds MBP weaker at P9 and stronger at P4 pos. 70 than susceptibility allele *03:01.
*03:01 shows a quite distinct P4 polar environment, respect to *14:01.
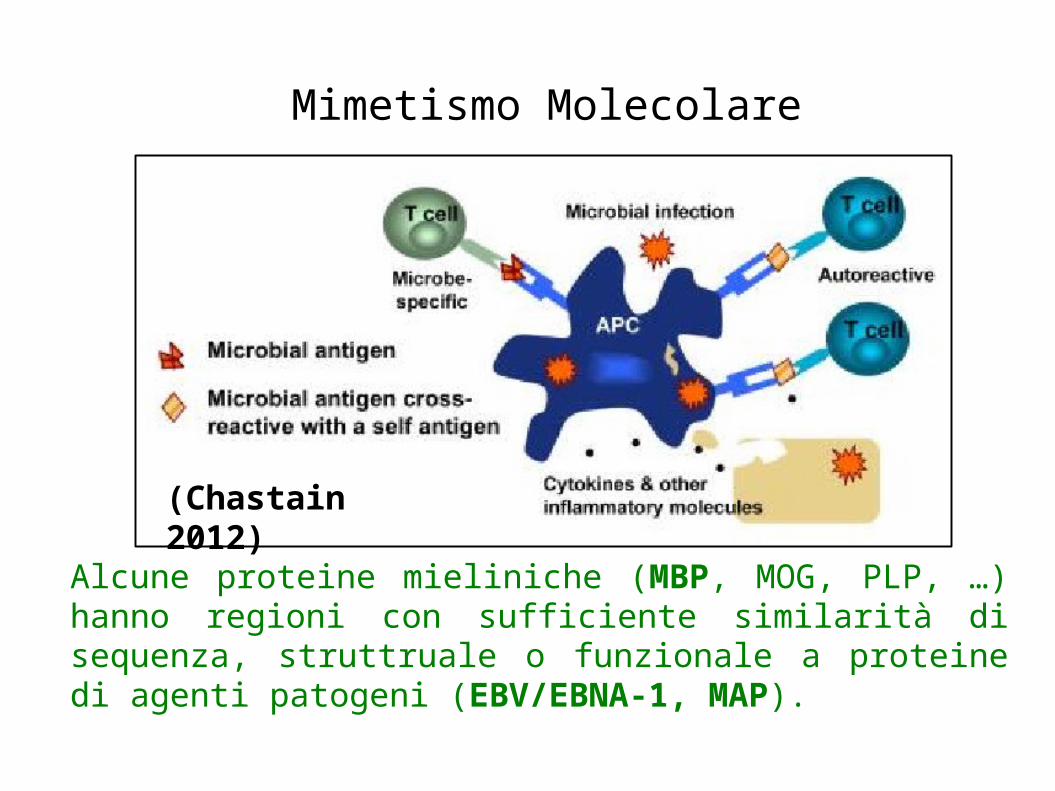
Mimetismo Molecolare
Alcune proteine mieliniche (MBP, MOG, PLP, …) hanno regioni con sufficiente similarità di sequenza, struttruale o funzionale a proteine di agenti patogeni (EBV/EBNA-1, MAP).
(Chastain 2012)
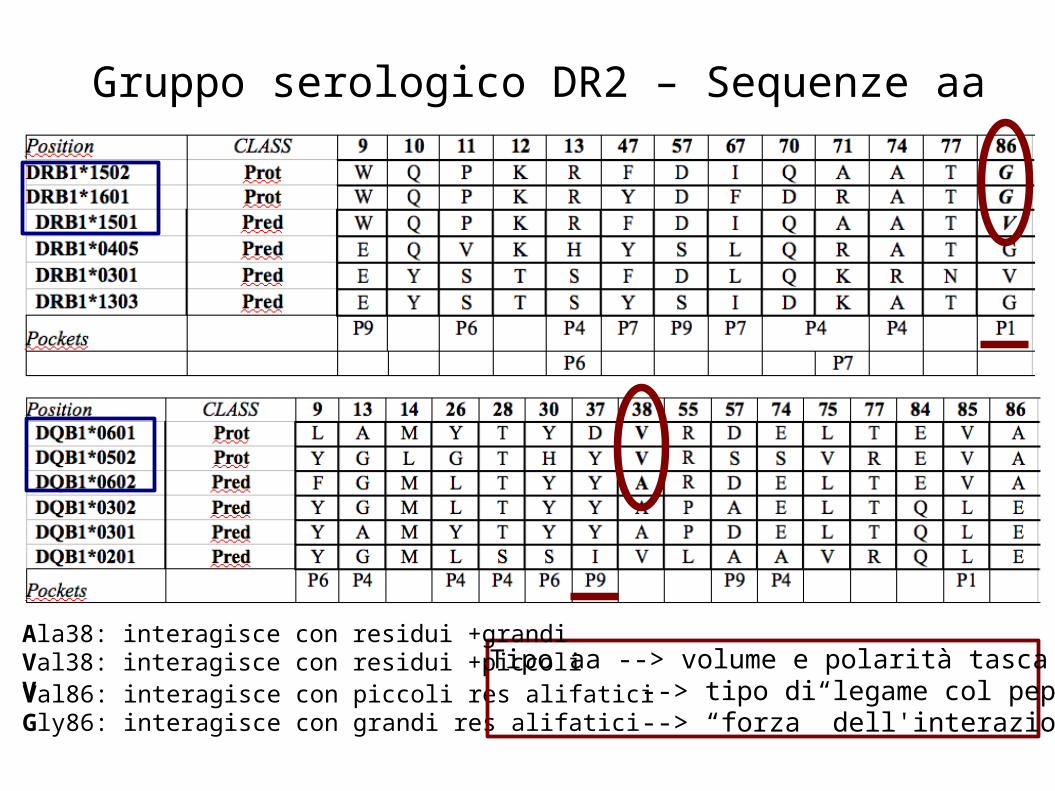
Gruppo serologico DR2 – Sequenze aa
Val86: interagisce con piccoli res alifaticiGly86: interagisce con grandi res alifatici
Ala38: interagisce con residui +grandiVal38: interagisce con residui +piccoliV
Tipo aa --> volume e polarità tasca --> tipo di legame col peptide--> “forza” dell'interazione col TCR
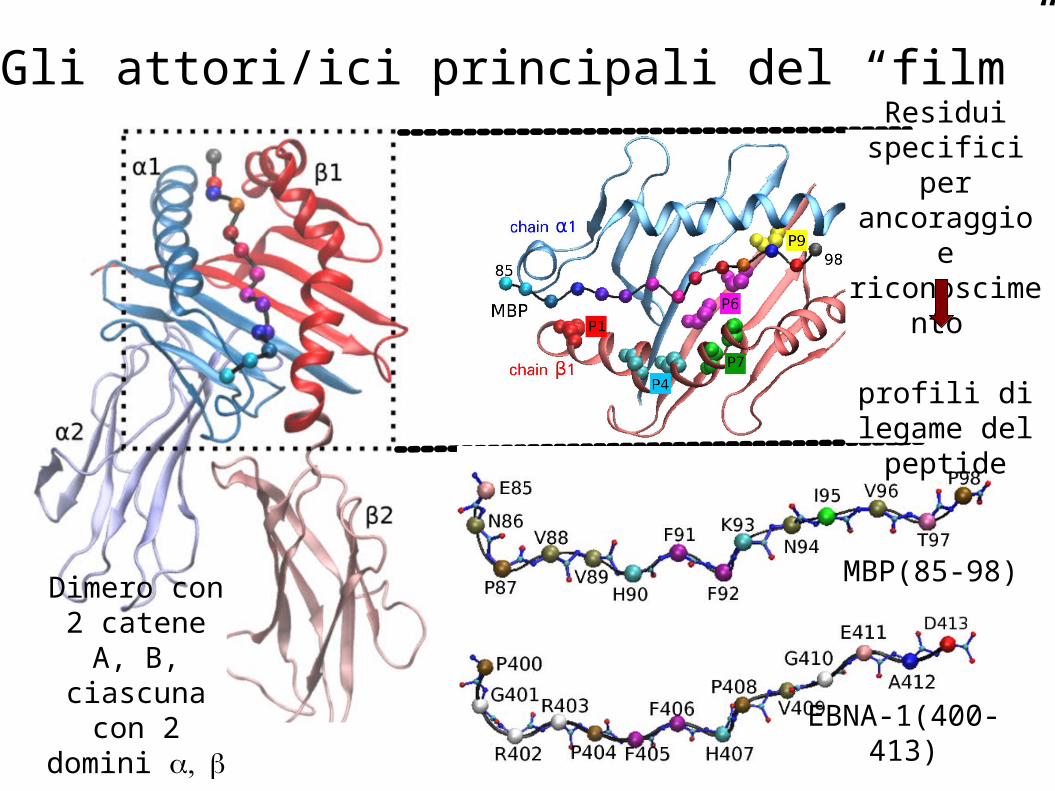
Gli attori/ici principali del “film”
MBP(85-98)
EBNA-1(400-413)
Dimero con 2 catene A, B, ciascuna
con 2 domini αβ
Residui specifici per ancoraggio e
riconoscimento
profili di legame del
peptide

![Le nuove TERAPIE DELLA SCLEROSI MULTIPLA [Sola lettura]](https://static.documenti.site/doc/80x56/61a136a9ad67dc289f5d8317/le-nuove-terapie-della-sclerosi-multipla-sola-lettura.jpg)















